13-2 Learning
Austin Lim, PhD and David Graykowski
Austin Lim, PhD (DePaul University) David Graykowski (DePaul University)
Editor: Alexandrina Guran, PhD (Universität Wien)
Think back to your favorite birthday party. Which of your friends were there? What did you do, where did you go, and did you have cake? Did you get gifts?
The ability to perform this task depends on our ability to create and recall memories. According to our current best understanding of the neuroscience of learning, the underlying biology of a memory mainly consists of subtle changes among synapses distributed across

Patient HMNeural Structures Involved in LearningCellular Mechanisms of LearningMolecular Mechanisms of Learning
Chapter 13 outline
Patient HM
One of the most influential case studies in the neuroscience of memory is the story of Patient HM. HM was born in 1926 in a small Connecticut town. He had a mostly regular childhood: taking family road trips, riding bicycles, and learning about American presidents in school.
In his childhood, HM began having severe seizures, possibly the result of a head injury. In his teenage years, he started having tonic-clonic seizures, the most severe form of seizures that produces a loss of consciousness and convulsions (extreme muscle contraction or
several brain areas. Our ability to learn new facts, recount the events of last week, or to perform new motor skills is the result of learning-induced neural plasticity. In this chapter, we will consider different aspects of learning and memory, starting from the behavioral level down to the molecular changes responsible for memory formation, as well as some disorders that disrupt healthy memory processes.
extension). In his early adulthood, he was having a tonic-clonic seizure monthly and several minor seizures daily, preventing him from working a normal job or living a normal life – despite taking a cocktail of anti-epileptic medications.
Neurosurgeon William Scoville proposed a “frankly experimental operation” to treat HM. It was known that most epilepsy originates in patches of neurons of the medial temporal lobe (MTL), and HM’s epilepsy was typical in this respect. Scoville suggested to surgically resect the MTL. In 1953, Scoville removed about
 Figure 13.1 Patient HM at 27 years old.
Figure 13.1 Patient HM at 27 years old.
8 cm of the MTL bilaterally, including part of the amygdala, and notably the hippocampus, the seahorse-shaped structure of the brain.
The surgery succeeded at its primary goal: HM’s seizures were less frequent and less severe. However, HM was left with a highly unusual and life-altering side effect: He was unable to create new discrete memories, a memory deficit called anterograde amnesia. For example, he could not remember what he had eaten for lunch just minutes after finishing the last bite. Despite being an avid fan of watching the news, HM couldn’t remember the names or the faces of different celebrities or public figures. It was as if he was permanently living in the present. (In contrast, retrograde amnesia affects the ability to successfully retrieve memory from one’s past.) However, despite his pervasive memory deficits, HM did not display any deficits in intelligence. His language and speech were unaffected, and word recall was excellent, as he loved completing crossword puzzles and often did so successfully late in life, with only the occasional spelling errors. He could learn to acquire new skills, such as keeping a pen still on a moving circular platform, or a tapping task (these skills
Hippocampus
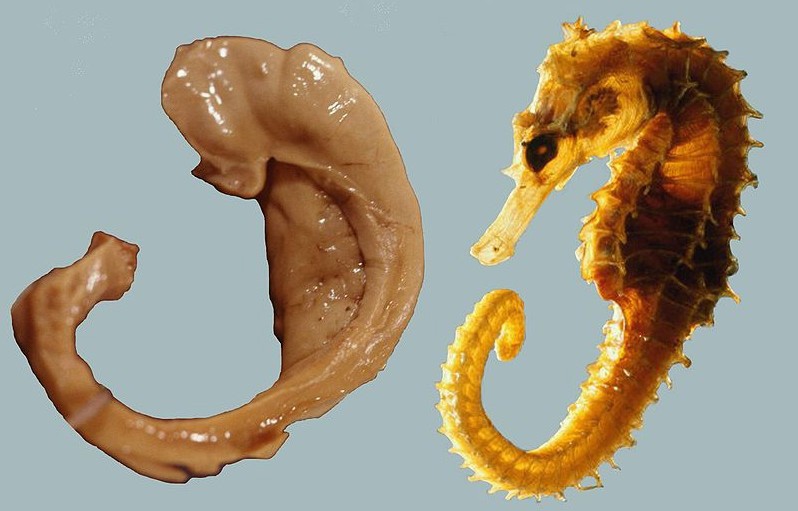
Figure 13.2 The location of the hippocampus in the medial temporal lobe (top). A dissected hippocampus and fornix (bottom left) looks like a seahorse (bottom right).
are different form of memory called procedural memory; see below). He was also capable of recalling things from his early childhood, such as geography facts he had learned in elementary school.
 Types of memories
Types of memories
The fact that HM’s MTL surgery disrupted some types of memories (e.g., memory for facts) while others were still intact (e.g., motor skills) inspired neuropsychiatrists to try to define the different forms of memory. Much of the research was led by Dr. Brenda Milner, who carried out several behavioral tests on HM to figure out what
types of memories are dependent on the intact MTL and which ones can function without MTL.
The most profound deficit was HM’s inability to create new declarative memories. Declarative memories, also called explicit memories, are the pieces of information that can be consciously declared or stated explicitly. Declarative memories are thought of as a “knowing what”. Declarative memories can be further subdivided into semantic memory and episodic memory.
Semantic memories are pieces of factual information. Some examples include:
- “Jupiter is the largest planet of our solar system.”
- “Rosalind Franklin discovered the double- helix structure of a DNA molecule.”
- “TheactorKeanuReevesplayedthe protagonist of the movie The Matrix.”
An episodic memory, sometimes also called an autobiographical memory, is the recollection of a discrete moment in a person’s life. It can be thought of as “mental-time travel”
– what was it like when. The following memories are examples of episodic memories:
- “When I got home, I put my wallet and phone on the table.”
- “I ordered pizza last night.”
- “In 2019, I went to see my favorite musicians perform live.”
Several tests concluded that HM had lost his ability to create new semantic memories. In one such study, HM was asked to determine if a word was made up or real. He was shown words with very old origins, such as “shepherd” or “butcher.” On these words, he performed as well as the control group. When he was shown words that are made up, such as “phlage” or “thweise”, he likewise performed as well as the controls. However, when shown words that were added into the dictionary after his 1953-surgery, such as “granola” or “jacuzzi,” he scored about 50% correct – consistent with guessing at random, as if he never acquired the knowledge that these words have a meaning.
HM was also unable to create autobiographical memories. When asked to recall one of his birthday celebrations as an adult, he wouldn’t be able to give any significant details about the event. Instead, his answers were often vague and generic.
One interesting observation was that HM’s memory about details from his childhood were still intact. The inability to recall memories from the past, in this case, from before HM´s surgery, is called retrograde amnesia. Patient HM’s retrograde amnesia was temporally graded, meaning that the farther back you examine, the more complete his memories were. Many of his memories for the two years before his
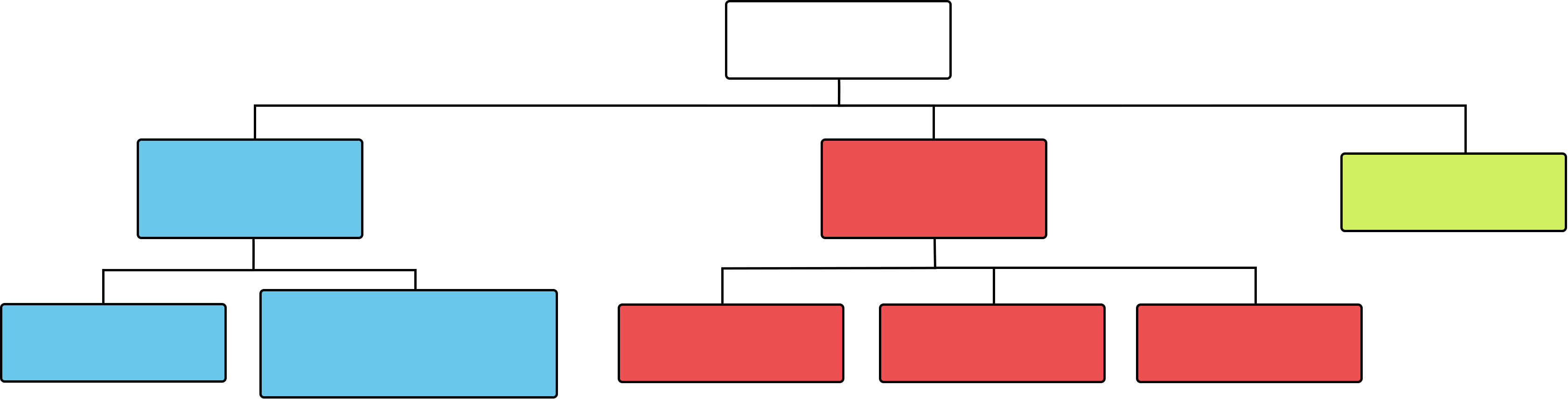 MemoryDeclarative (explicit)Procedural (implicit)WorkingSemanticEpisodic (autobiographical)KinestheticPrimingAssociative
MemoryDeclarative (explicit)Procedural (implicit)WorkingSemanticEpisodic (autobiographical)KinestheticPrimingAssociative
Figure 13.3 Summary diagram of some of the major subtypes of memory.
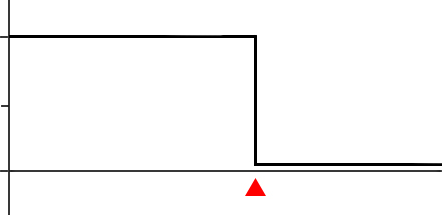 Anterograde amnesiaTimeLesion
Anterograde amnesiaTimeLesion
Memories retained
100%
50%
0%
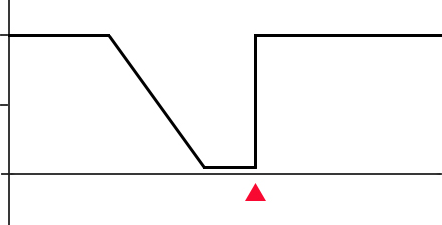 Retrograde amnesiaLesionTime
Retrograde amnesiaLesionTime
Memories retained
100%
50%
0%
Figure 13.4 In anterograde amnesia, a person is unable to create new memories following a lesion (top). In temporally-graded retrograde amnesia, older memories are better retained while recent memories are more likely lost.
surgery were completely lost, but memories from his youth and teenage years were intact as much as healthy individuals (there is contention about this observation, because HM was taking several anti-epileptic drugs, which may have impacted memory formation.) From this observation, memory researchers concluded that the MTL functions as short-term storage site for memories, but after some years, those memories get relocated to other brain areas outside of the MTL. Currently, the scientific evidence suggests that memories are distributed across several networks of cortical and subcortical brain areas.
While HM lost the ability to create new declarative memories, he was still able to maintain a different class of memories, called procedural memories (or implicit memories). They are unconscious memories, and can’t be explicitly
stated. These can be thought of as “knowing
how”. Some examples of procedural memories include, for example, performance of a series of motor actions without conscious thought such as an experienced musician playing a simple scale (sometimes commonly called “muscle memory”, even though the muscles do not store any actual memory!), or a priming effect (such as when a person sees pictures of bananas, they are more likely to answer the fill-in-the-blank prompt “b _ _
_ _ _” with “banana”, whereas other people might guess “bubble” or “badger”).
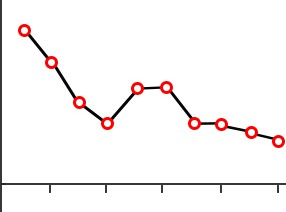 Day 1Trials
Day 1Trials
Errors
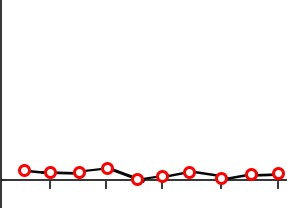
The original test of procedural memory conducted by Dr. Brenda Milner was called the mirror tracing task. In this test, the patient is told to draw a third star in between the two stars as quickly as possible without making any mistakes. The challenge is that the tracing is to be done while watching their hand and the star in their reflection in a mirror. Because of these unusual circumstances, completing this task is difficult. But over multiple days of practice, people become
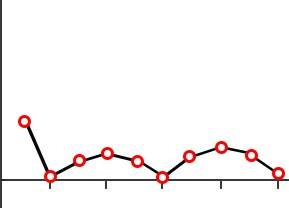 Day 2Trials
Day 2Trials
 Day 3Trials
Day 3Trials
Figure 13.5 Patient HM performed poorly on the mirror tracing task (top), but improved at the task over time despite having no memory of performing the task (bottom).
better at this mirror tracing task, completing it faster with fewer errors. Improvement on this task indicates that a person is learning or gaining some memory about how to better perform the task.
After practicing this mirror tracing task, HM was able to finish drawing the star about ten times faster than when he first began. He improved his performance within each day’s worth of training, and he also improved day-to-day. There is evidence that he maintained these skills up to one year later, despite not having regular training on this task. Surprisingly, each day Milner examined HM, she would need to reintroduce herself since he forgot who she was. She also had to re-explain what HM was supposed to do in the mirror tracing task. Hence, while HM was unable to form declarative memory about the experiment or the people involved, learning of the procedural memories and motor actions involved in this task remained intact.
Another type of procedural memory is an associative memory. Associative memories are the types of information that we learn through traditional Pavlovian conditioning. For example, recall the classic Nobel prize-winning experiment in physiology conducted by Ivan Pavlov in the late 1800s. Normally, the presentation of dog food, an unconditioned stimulus (US), causes a dog to salivate, a naturally happening behavior, called the unconditioned response (UR). Dogs are not particularly interested in the sound of a whistle: this neutral stimulus will produce a minor response, such as a head turn and attentional shift towards the origin of the sound, but not much
dogs begin to salivate, a conditioned response (CR), independent of any food being presented.
Separate from declarative or procedural memories, a different form of memory called working memory was tested in HM. Working memory involves processes of storing information temporarily while simultaneously manipulating those pieces of information. This type of memory

 Food: Unconditioned stimulus (US)Bell: Neutral stimulusPairing of bell with foodBell: Conditioned stimulus (CS)
Food: Unconditioned stimulus (US)Bell: Neutral stimulusPairing of bell with foodBell: Conditioned stimulus (CS)
StimulusResponse
Salivation: Unconditioned response (UR)
No response
Salivation
Salivation: Conditioned response (CR)
more than that. However, when this stimulus is
repeatedly paired with the presentation of food, dogs quickly learn to associate that the whistle signals food. After multiple pairings, upon hearing the whistle, a conditioned stimulus (CS), the
Figure 13.6 A CR after exposure to a CS, such as in classical Pavlovian conditioning, is an example of an associative memory, one type of procedural memory.
can be thought of as a “short-term memory on overdrive.” Although HM struggled with working memory immediately after his surgery, several
Trial 1:Prompt Expected response
4 8 2 6
6 2 8 4
years later HM performed as well as age-matched
Trial 2:
Prompt
2 8 7 9 1
control patients on these tasks.
For example, a test of working memory
Expected response
1 9 7 8 2
is the digit span test, where a person is given
Trial 3:
Prompt
8 4 1 1 8 6
a series of numbers to remember, then they are asked to repeat the numbers in reverse order.
Expected response
6 8 1 1 4 8
After successfully completing this task, a different series of numbers, this time one digit longer, is presented to the patient until they first start making errors in recall. A related task is called the Corsi block tapping test, where an experimenter sets up several blocks on a table. The experimenter
PromptExpected response
 4132
4132
 1423
1423
then taps a series of blocks in a specific order,
then the subject is asked to tap on the blocks in reverse order. As with the digit-span test, the experimenter then makes the series of blocks longer until they make mistakes in the tapping.
Patient HM died in 2008 at age 82 of respiratory failure. His name was Henry Molaison.
Figure 13.7 The digit span test (top) and the Corsi block tapping test (bottom) are measures of working memory.
Neural Structures Involved in
The Hippocampus (HPC)
 Schaffer collateralsCA1CA3DentateMossygyrus fibersPerforant pathwayEntorhinal cortex
Schaffer collateralsCA1CA3DentateMossygyrus fibersPerforant pathwayEntorhinal cortex
The hippocampus (HPC), meaning “seahorse” in Greek, was named based on its morphology. The HPC is located along the ventral and medial surface of the brain. The HPC is one of the critical structures of the limbic system, a series of subcortical brain structures that are involved in several different complex behaviors, such as emotions and memory. The limbic system is an evolutionarily ancient brain network.
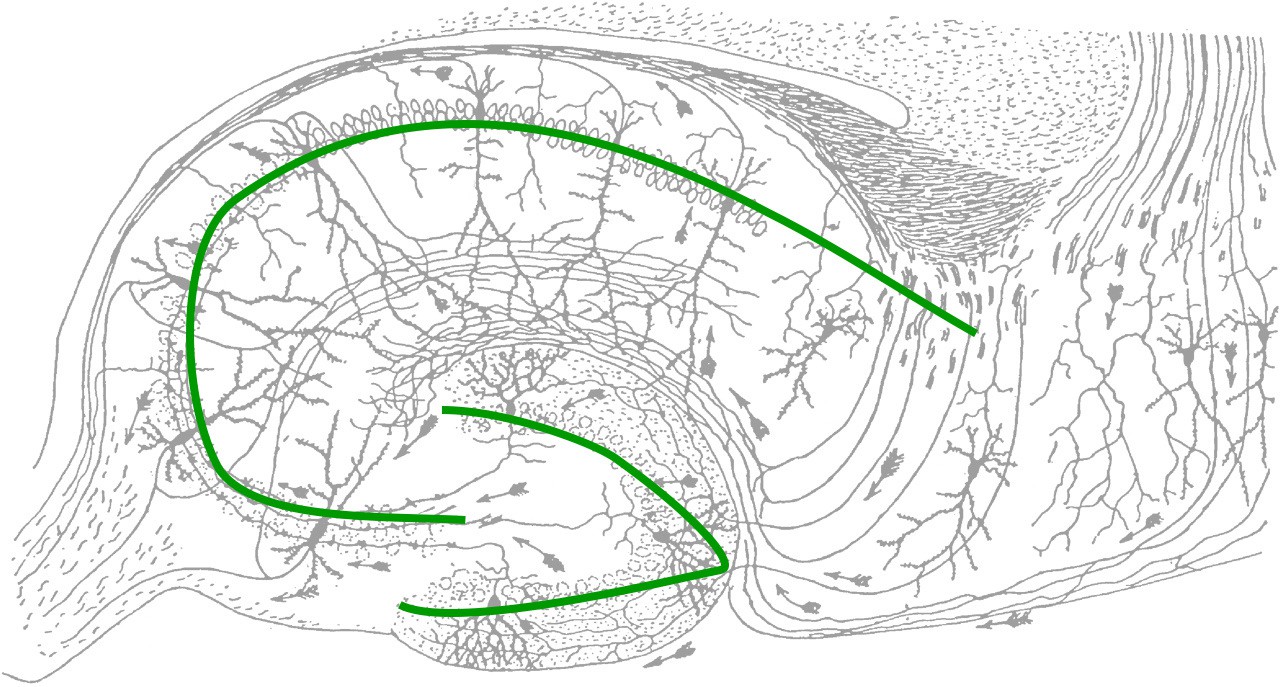
The synaptic connectivity of the hippocampus is very well characterized. Hippocampal synaptic connectivity was first described by Ramon y Cajal, and is made up of three main synaptic connections; sometimes
called the trisynaptic circuit. First, the axonal
outputs of layers 2 and 3 from the entorhinal cortex make up the inputs into the HPC. This white matter signaling tract is called the perforant pathway, and they synapse onto the granule cells of the dentate gyrus. These neurons send axons, called mossy fibers, to the pyramidal cells of the Cornu ammonis (CA) 3 region of the HPC. The axonal projections from here, called Schaffer collaterals, project into CA1, which are the neurons that make up the output of the hippocampus. These outputs project out to layer
5 and 6 of entorhinal cortex. While the three main neuronal projections are glutamatergic, the trisynaptic circuit is modulated by GABA, acetylcholine, norepinephrine, and serotonin.
The HPC is involved in spatial memories, memories involved in navigation of our surroundings and the creation of a mental map of our world. Spatial memories are developed when we enter a new building for the first time, and we search for a new classroom. We also use our
Figure 13.8 The circuitry of the hippocampus as illustrated by Ramon y Cajal (top) and as a schematic diagram (bottom). Structures are labeled in red and communication pathways are in black.
spatial memory whenever we are walking around campus, making our way from one building to another, thinking about the streets you’d need to cross or the buildings you can cut through. While the volume of the hippocampus is not a reliable indicator of the strength of a healthy person’s spatial memory, injury to the hippocampus causes deficits in spatial memory.
To test spatial memory behaviorally in non- human animals, one test that is regularly used in rodents is called the Morris water maze. In this test, a shallow pool is filled with an opaque liquid, making it difficult to see through. Hidden somewhere in this pool is a clear plexiglass
platform, surrounding the pool are different
environmental cues that can be seen from the surface of the water, such as different shapes or colors. The water is deep enough that when a rodent is put into the Morris water maze, they have to swim to stay afloat. The rodents swim around aimlessly until they find the platform, the time it takes for this to happen is recorded, and the trial ends. Over time, the animals learn that the platform is located near certain navigational cues, and on future trials, the animals spend more time near those cues, and the latency to
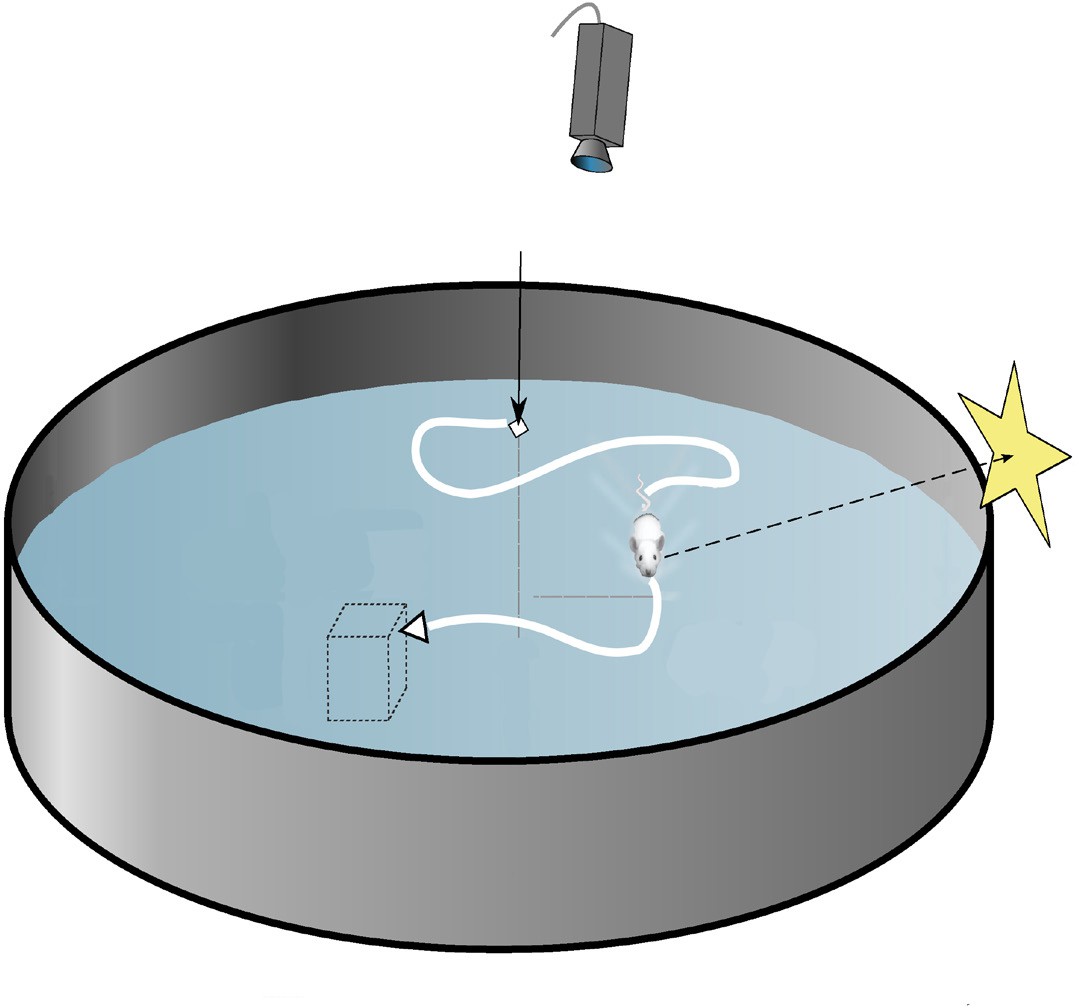 CameraStarting locationEnvironmentalcuesHidden platform
CameraStarting locationEnvironmentalcuesHidden platform

Figure 13.9 The Morris water maze (top) and the radial arm maze (bottom) are behavioral tests to assess spatial memory.
find the platform decreases. When the HPC is surgically removed from rodents or inactivated, they perform poorly in the Morris water maze.
Another non-human behavioral test used to assess the capacity for learning navigational cues is the radial arm maze. In this test, a rodent is placed on a circular platform. Extending from this platform are eight or more “arms”, at the end of each is a small dish. In one of the dishes is a morsel of food (“rewarded arm”), while the other dishes contain nothing (“non-rewarded arms”). The maze is designed so that the food cannot be seen from the end of each arm, so the animal must return to the starting platform before exploring another arm. The number of entries into a non-rewarded arms is counted as an error. Over time, the animals make fewer errors as they learn which arm is rewarded and which ones are not. Alzheimer’s disease model organisms perform poorly on this task.
Based on the deficits seen in Patient HM and other experimental manipulations of the HPC, we conclude that the HPC is strongly implicated in the process of declarative memories and spatial navigation. Since some of HM’s memory functions were still intact, such as procedural memories and working memory, it is believed that these functions are independent of HPC function.
The Amygdala
The amygdala is another limbic system structure found in the medial temporal lobe adjacent to the HPC. Amygdala comes from the Greek word meaning “almond,” which roughly describes its shape. While the amygdala is often spoken of as a single structure, it is more accurately divided into several subnuclei, each with different cell populations and functions. One broad division distinguishes the basolateral amygdala (BLA) versus the central nucleus of
amygdala (CeA): The BLA contributes to both fear memories and reward processing, while the CeA contributes more to the physiological response in emotions as well the perception of emotion.
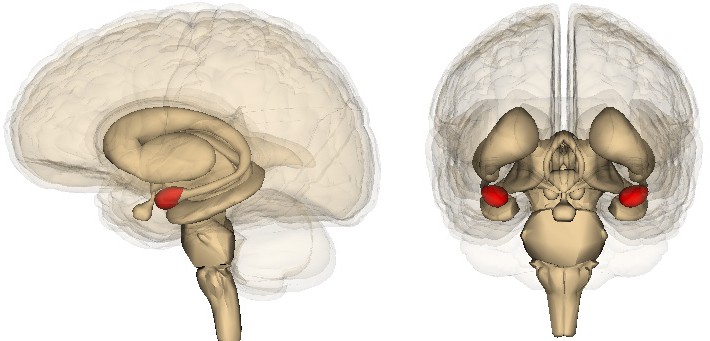
Figure 13.10 The amygdala are temporal lobe structures that contribute to the salience of emotional stimuli.
The amygdala is strongly involved with the formation and storage of emotional memories, memories or associations that have a strong emotional connection. Both positive and negative emotional states are represented here. For example, a whiff of grandmother’s cooking may cause you to reminisce back to a fun childhood summer. Alternatively, the smell of vomit may elicit the unpleasant emotions and nausea associated with a nasty food poisoning incident.
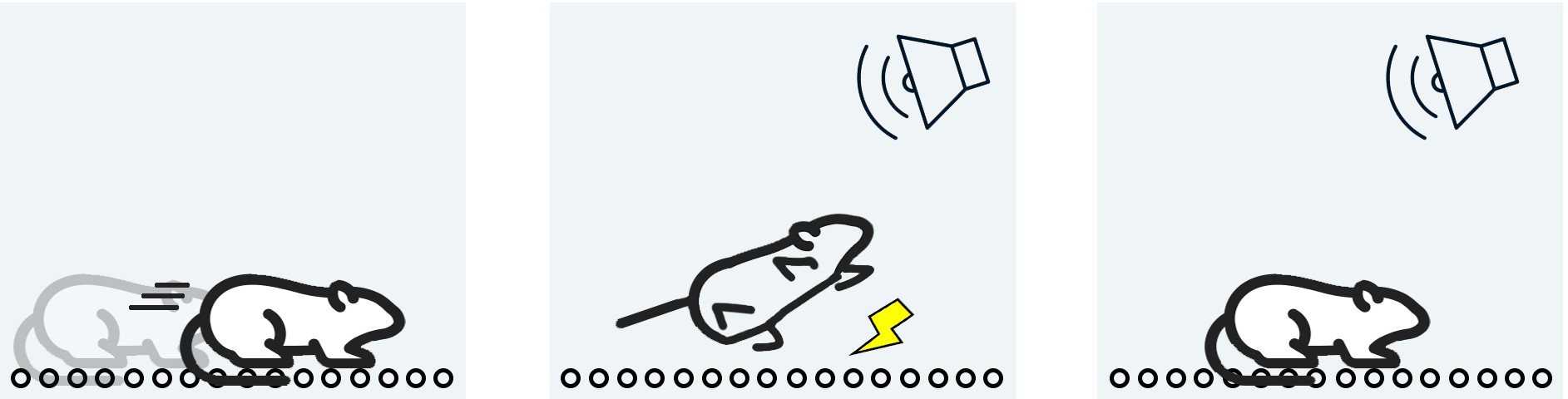
One non-human test of emotional memory is the foot-shock paradigm, a form of
fear conditioning. This test involves putting a rodent into a chamber with floors made of metal rods, which are connected to an electric current generator. The metal rods can deliver a non- lethal but painful electric shock to the rodent’s foot. In this learning paradigm, a combination of sound and light cues is presented to the animal. Shortly after, the painful foot shock is delivered. If the animal learns that the cues are associated with the negative painful memory, they exhibit freezing after exposure to the cues. Amygdala lesions prevent the animal from freezing, while hippocampal lesions have no effect on this emotional learning. Changing cellular signaling in the amygdala alters the learning of fear conditioning. The foot-shock paradigm is often used as a non-human model of post-traumatic stress disorder.
Inferotemporal cortex (IT)
 Structuresoftheinferotemporalcortex(IT) are part of the ventral stream of visual perception (chapter 7). The IT stores some components of visual memory. We use these functions when we see a classmate outside the classroom and recognize them from our Introduction to Neuroscience class, or when we see a parody of a famous painting and recognize the similarities to the original work. A simple behavioral task to assess visual memory would start by
Structuresoftheinferotemporalcortex(IT) are part of the ventral stream of visual perception (chapter 7). The IT stores some components of visual memory. We use these functions when we see a classmate outside the classroom and recognize them from our Introduction to Neuroscience class, or when we see a parody of a famous painting and recognize the similarities to the original work. A simple behavioral task to assess visual memory would start by
viewing a series of abstract shapes, and when a shape appears that you have already seen, you push on a button. The human capacity for visual memory is massive: After viewing
Figure 13.11 In the fear conditioning paradigm, a rodent is put
into a room with medal rods as the floor (left). Then, a sound tone is repeatedly paired with a foot shock (middle). When the sound is played again, the rodent may exhibit freezing behavior (right).
10,000 images for a few seconds apiece, people were able to identify a previously seen image successfully about 83% of the time.

Figure 13.12 The inferotemporal cortex is one of the signaling pathways important for visual memories.
A bit more specifically, one part of the IT is the fusiform gyrus, which has been previously described in the context of facial recognition. People with prosopagnosia, a visual perceptual disorder affecting the fusiform gyrus, can perceive the different parts of a person’s face, but have a difficult time putting the whole picture together and matching those features to a specific person. For facial recognition to be accurate, there must be some memory that allows for a person to
match those facial features with someone they have seen before, which is a memory related process.
The parahippocampal place area (PPA), also found in IT, contributes to visual memories associated with locations and environmental scenes. Imaging studies have demonstrated that activity of the PPA increases specifically when people view place-related images, including scenic landscapes like mountains, man-made structures like campus buildings, or the interiors of rooms, both furnished and completely empty. To serve as control stimuli, viewing faces or objects does not increase the activity of the PPA.
Prefrontal Cortex (PFC)
Aspartof the frontal lobe, the PFC is involved in high order decision making and personality. In the context of memory, neural circuits in PFC are important for short-term and working memory. Patients with injuries to their prefrontal cortex after stroke, tumors or aneurysm, performed worse on a variety of working memory tasks such as the digit span test. Additionally, people with frontotemporal dementia, a neurodegenerative
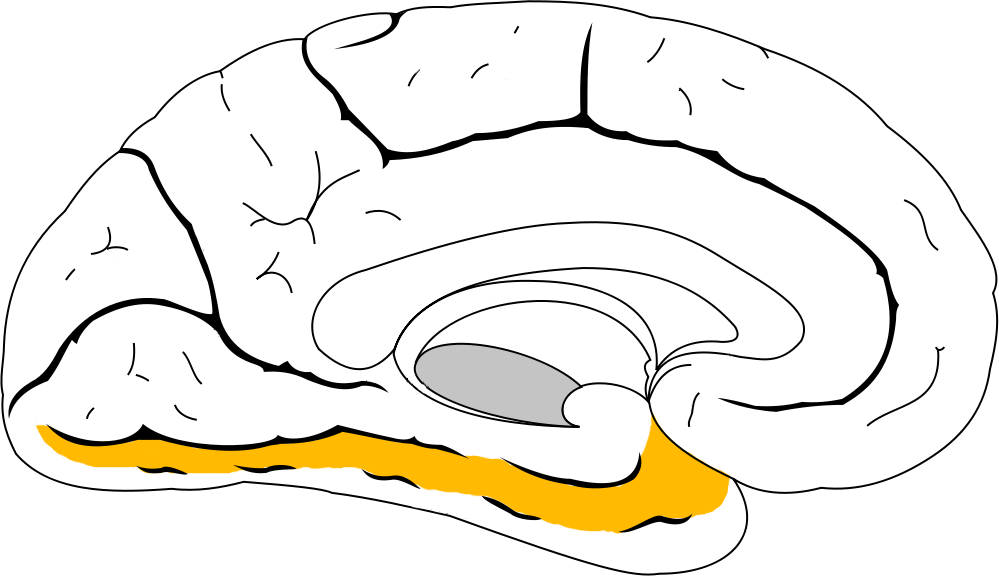
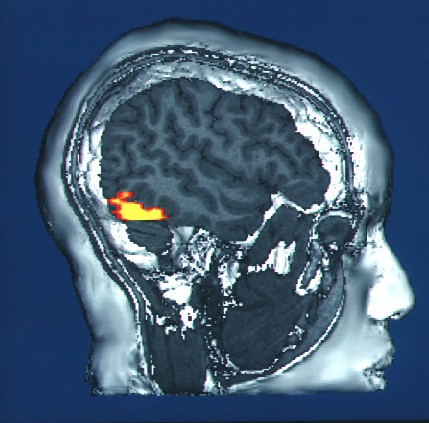

Figure 13.13 The inferotemporal cortex (left, sagittal view) is part of the ventral stream of visual perception, and is likely one site of visual information storage. Within the IT is the fusiform gyrus (middle), which is specifically activated strongly in imaging studies when a person is shown with a facial stimulus (right).
disorder characterized by a degradation of the frontal lobe, often have difficulty with working memory.
The PFC also has strong projections with the hippocampus, and these circuits are likely also involved in the formation of hippocampal- dependent memories.
Striatum
The striatum is a structure of the basal ganglia, a series of brain structures that contribute to behaviors such as motor activity (chapter 10) and procedural memories. The striatum likely holds memories involved in habits. Habitual behaviors help us preserve cognitive bandwidth, reducing the “mental energy” that is used during repetitive task performance. The downside of habits is that reliance on habitual responding can limit behavioral flexibility, and cause a person to act in a suboptimal manner, perhaps behaving in a way that led to a positive outcome in a previous
set of circumstances without incorporating and evaluating the present circumstances.
Habitual action performance is likely related to a variety of neuropsychiatric disorders. Obsessive compulsive disorder (OCD), for example, is characterized by the presence of recurring, intrusive thoughts, which can lead to repetitive actions. Commonly observed is the thought that one’s hands are unclean, which leads to repeated handwashing.
A rodent behavioral test of habitual activity is the observation of self-grooming, a natural and healthy series of stereotyped actions that consists of licking the paws and moving them through the fur of the nose, caudally down the body. Mouse models of OCD show excessive self-grooming to the point where they pull their fur out and paw their skin to the point of injury.
Drug addiction is also a striatal disorder. Compulsive drug use is often associated with a series of habitual motor actions that happen
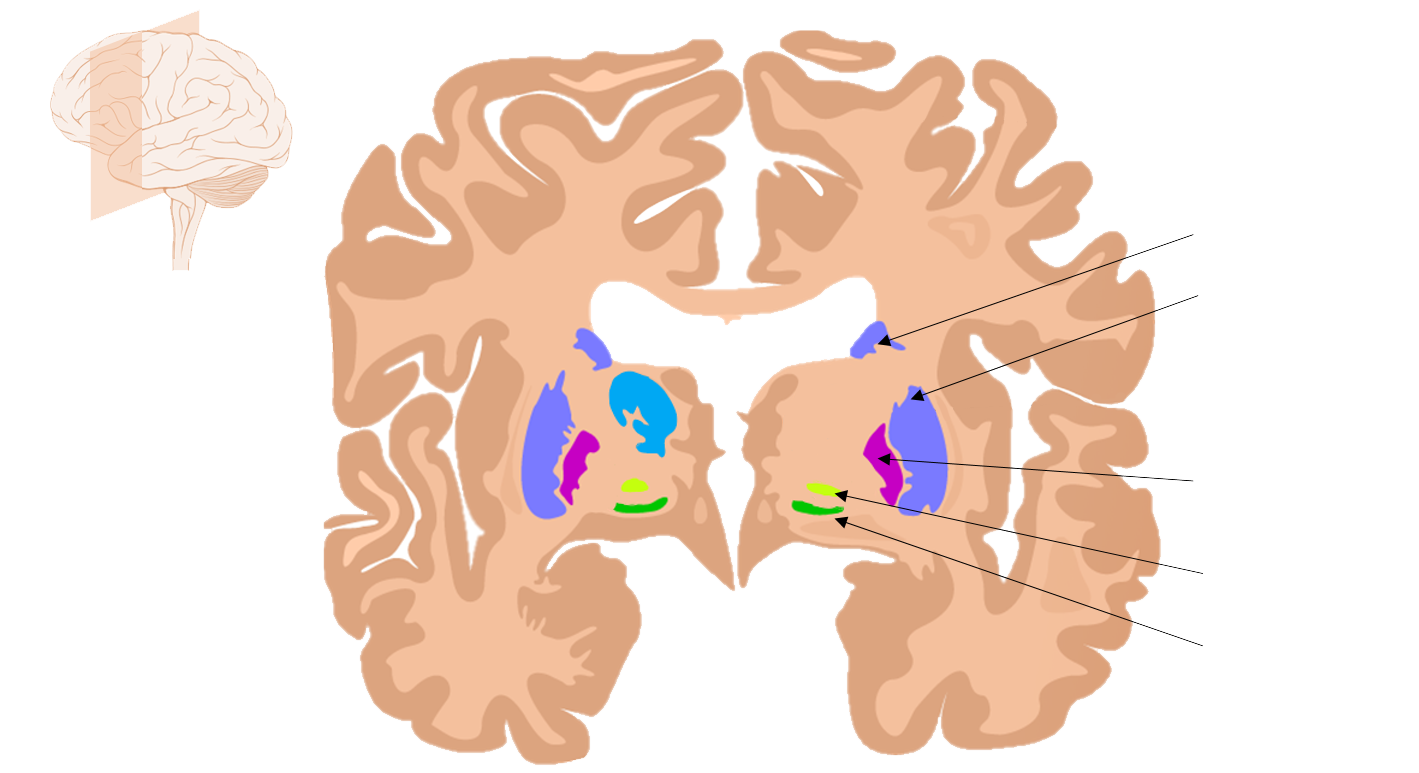 Caudate Putamen
Caudate Putamen
Globus pallidus
Striatum
Subthalamic nucleus Substantia nigra
Figure 13.14 Several subcortical brain areas make up the basal ganglia.
HypermnesiaSolomon Shereshevsky was one of a handful of rare, clinically documented cases of hypermnesia, the capacity to recall nearly any memory with perfect precision, even after several years. Remarkably, he could “easily remember any number of words and digits, equally easily he memorizes whole pages from books on any subject and in any language and for a quite long time at that. Shereshevsky can accurately quote anything he was told ten or twelve years ago.” He received recognition in 1968 when his psychologistA. R. Luria published a case study in “The Mind of a Mnemonist.”Today, we would describe Shereshevsky as being autistic with strong multimodal synesthesia. Also of note, he had significant deficits in executive function, difficulty with recognizing faces, and could not interpret abstract ideas.
before a person experiences the drug effect. For example, in tobacco use disorder, people will perform an orchestrated series of actions, including opening a pack of cigarettes, flicking the lighter, withdrawing the cigarette and taking a deep inhalation. Some of these behaviors are likely stored across striatal circuits (chapter 11).
Cerebellum
The cerebellum is the phylogenetically ancient structure found posterior and ventral to the cerebrum, and functions generally to help with motor functions (chapter 10). The cerebellum is involved in procedural memories, particularly the performance of motor abilities. Learning new motor skills likely requires changes in the circuit strength of cerebellar neurons.
This list of brain structures involved in memory is certainly not exclusive. For example, the orbitofrontal cortex plays a role in positive emotional memories, and sensory cortices are important for the memories related to the specific stimuli that are processed in those areas. A single memory could likely be stored in several brain areas, much like a mosaic.
Cellular Mechanisms of Learning
In the late 1800s, around the time when Golgi and Ramon y Cajal were engaged in intense debate about the organization of the nervous system, many neuroscientists came to a strange observation: the weight of the brain increases dramatically over the first 10 years of life, but not much more after that. Even though we learn lots of new facts and make lots of new memories in adulthood, the brain itself doesn’t grow in size. So how is it possible to store new knowledge if the brain is not making many new neurons?
Most likely, new pieces of information are held in the connections between cells, not just in the cells themselves. If our estimate of 150 trillion synapses per adult brain is correct, then it is possible that we could store all the knowledge and memories that we collect over our lifetime through some combination of activity across certain connections.
The activity at the cellular level is believed to take place on at least three different levels enabling us to build, store, and retrieve memories.
- Encoding refers to the ability for brain circuits to store some piece of information. In real life, you are presented with countless stimuli simultaneously. Imagine walking down a busy street, and think of the number of different sights, smells, and tactile stimuli you experience. Storing memories is an energetically costly process, and we are limited in the fact that all of our sensory inputs cannot possibly get encoded. Instead, evolution has preferred to encode stimuli that are most salient pieces of information, such as perceptual cues associated with predators. Alternatively, information that we pay strong attention to can get encoded more strongly, like
when we repeat a phone number to ourselves until we have a chance to write it down. It is also easier to encode novel information that “builds” on previous bits of knowledge, or information that is closely related to other well-established information, which is why analogies are such an effective way to learn new facts.
- The process that enables memory storing is called consolidation, which makes the memory more permanent. In 1949, an early neuropsychologist, Donald O. Hebb, offered an explanation for how changes in synapses could possibly lead to a phenomenon as complex as learning. His theory, published in his text The Organization of Behavior, can be summed up in the phrase:
“Cells that fire together, wire together.”
In Hebb’s framework, repeated activity at a synapse within a circuit of neurons acts as a reinforcer signal that strengthens this synapse for future communication, making the next incoming signal more robust. Hebb also implies the inverse is true: when cells do not fire together, they weaken their connection. Through fine tuning of synaptic connections, some strengthening and others weakening, a lifetime of memories can be stored across a wide distribution of neurons. After a memory has been created, the specific circuit of neurons that represent that piece of information is called a memory trace or an engram.
A cellular process called reverberation is thought to be the mechanism that allows for consolidation. Reverberation is the process by which networks of neurons fire repeatedly. Each
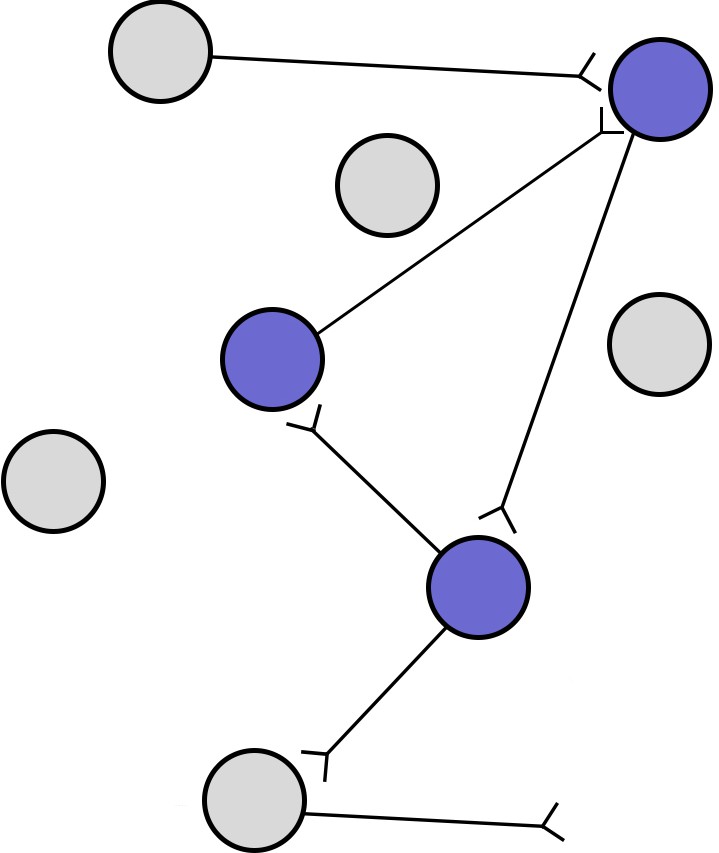 InputTo output
InputTo output
Figure 13.15 A reverberating circuit (purple) is a series of neurons that are activated repeatedly with the activity of a positive feedback circuit.
time that circuit is activated, the strength of the network is increased, meaning that it becomes easier for that circuit to be activated in the future. Throughout the process of consolidation, memory traces are thought to become more represented in the neocortex and less in the subcorticalstructureslikehippocampusor amygdala. Recall Patient HM’s temporally graded retrograde amnesia, where he had lost declarative memories in the two years leading up to this surgery, and yet his memories from the long past were maintained. This finding suggests that some aspects of declarative memory consolidation depends at least partially on medial temporal lobe and HPC for a period of time, maybe up to two years, before those memories get stored in the cortex more permanently. However, we cannot conclude how long consolidation takes in healthy humans based on findings from HM, who had pervasive and frequent seizures, which might
have negatively impacted memory consolidation before the surgery.
Consolidation seems to occur predominantly during sleep. Specifically, declarative memory is enhanced during non-REM sleep, while procedural memory is enhanced during REM sleep. Studies investigating sleep consolidation are often done by letting participants learn or perform behavioral tasks, after which they are deprived of a specific phase of sleep. To wake participants at the right moment, researchers commonly use EEG signatures, which are unique for different phases of sleep: sleep deprivation early in the night denies non-REM slow wave sleep, while late-sleep deprivation decreases time spent in REM sleep. Some speculate that the hallucinations we sometimes experience during dreaming are a consequence of consolidation processes, but it is inconclusive as to what role dreaming plays in memory formation.
- Finally, for the stored memories to be recalled, a cellular process called retrieval happens which brings back the specific engram. Retrieval happens for both declarative and procedural memories.
The writing of memories can be differentiated from the retrieval of memories using specific priming-related behavioral tests. One example is a vocabulary recall test. Consider if you were given a list of 50 words to memorize, words that belong to a handful of different conceptual categories (such as cinnamon, pepper, or curry, which fall under the category “food flavors”). When asked to write down as many words as possible from the list using your memory, a test called free-recall, you may be able to successfully remember about a third of them. However, if you were prompted with the category titles, a related test called cued-recall,
you would perform much better at retrieving those memories, possibly recalling up to 75% of the words. The fact that cued-recall scores are often higher than free-recall scores indicate that there is a distinction between the encoding
/ consolidation of memories and the retrieval of memories.
Retrieval is not a passive function. When an engram is retrieved, it is reconsolidated, which is an act similar to replaying the activity of the circuit. During this reconsolidation, it is possible that some aspects of the memory are emphasized, while others are lost. This is likely the main reason why we experience false memories, memories that are not true to reality
– one reason why eyewitness testimonies are notoriously unreliable. We may imagine our good memories as better than they actually were, while simultaneously dampening the negative aspects of those memories. A dysregulation of this reconsolidation process could lead to the symptoms seen in post-traumatic stress disorder, where the negative emotional components of a particular memory are exaggerated rather than being blunted.
Special populations of neurons
An individual memory is likely distributed widely across several different parts of the brain. However, there are a few special populations of neurons mainly in the MTL that contribute to highly specific types of memories.
Place cells
Place cells are a special population of pyramidal cells of the hippocampus. These neurons increase their firing activity when the animal is in a particular location in an environment, indicating that they contribute strongly to location and navigational memory. There is no apparent
topographical arrangement of these place cells, meaning that adjacent areas of an environment do not necessarily activate adjacent hippocampal place cells. The place cells, when firing at the right times, help the animal create a spatial map of their surroundings.
Grid cells
 Grid cells are located in the entorhinal cortex, the main input structure to the HPC. Closely related to the place cells described above, grid cells increase their firing properties
Grid cells are located in the entorhinal cortex, the main input structure to the HPC. Closely related to the place cells described above, grid cells increase their firing properties
Neuron 1
Neuron 2
Neuron 3
Neuron 4
Figure 13.16 A rat is tracked (left, gray) as they move through an open field. Individual grid neurons spike when the rat passes through particular areas of the open field (left, red). Heat map (right) showing high neuronal activity in warm colors (red and yellow) with low activity in cool colors (blue).
periodically when an animal is at an intersection of a “grid” in a wide-open, previously-explored environment. The grid itself is roughly hexagonal, and spans the whole environment an animal is in. The overlap of multiple grids gives the animal an idea of the surroundings. The scientific description of grid cells earned three scientists a Nobel Prize in Physiology or Medicine in 2014.
Jennifer Aniston neurons
The “Jennifer Aniston” neurons, also called concept cells, are a series of cortical neurons in the temporal lobe that increase their firing exclusively in response to highly-specific stimuli, such as the idea of Jennifer Aniston, Halle Berry,
or the Tower of Pisa. These concept cells respond to much more than just pictures: for example, a Luke Skywalker neuron that responded to a picture of young Mark Hamill (the actor who played Luke Skywalker) will also respond to text that reads “LUKE SKYWALKER” and the sound of a person saying “Luke Skywalker”. Although this particular neuron probably won’t fire in response to pictures of athlete Manu Ginobili or actress Marilyn Monroe, the neuron might fire in response to pictures of Yoda or Darth Vader, indicating that the neuron may encode an even broader concept, such as “Star Wars characters” or “Jedi”, or is a part of a network that encodes concepts related to the Star Wars franchise.






Figure 13.17 Concept cells change their firing pattern in response to the presentation of highly specific stimuli, such as the character Luke Skywalker (portrayed by actor Mark Hamill; images 1, 3, and 5). Visually similar but conceptually different stimuli (male brunette actors appearing in film), like pictures of Leonardo diCaprio (image 2) or Keanu Reeves (image 4), fail to induce changes in firing. However, visually distinct but conceptually-related stimuli, like the picture of Yoda (a related character from the same series of films; image 6) may also drive the concept cells to fire.
Molecular Mechanisms of Learning
Zooming in beyond the level of anatomy, the substrates of learning can be found at the level of synapses. Synapses change in a phenomenon called plasticity. The word “plasticity” refers to a change in synaptic strength, which may be an increase or a decrease. This change may persist for minutes, hours, days, or in some cases, even a whole lifetime. When synaptic strength is increased and remains elevated, we call this long- term potentiation (LTP). A prolonged weakness of a synapse is called long-term depression (LTD). In our current limited understanding of plasticity, both phenomena are important for a healthy brain, and neither one is always good or always bad.
It is also important to clarify that both excitatory synapses and inhibitory synapses can be subject to either LTP or LTD.
Long-term potentiation
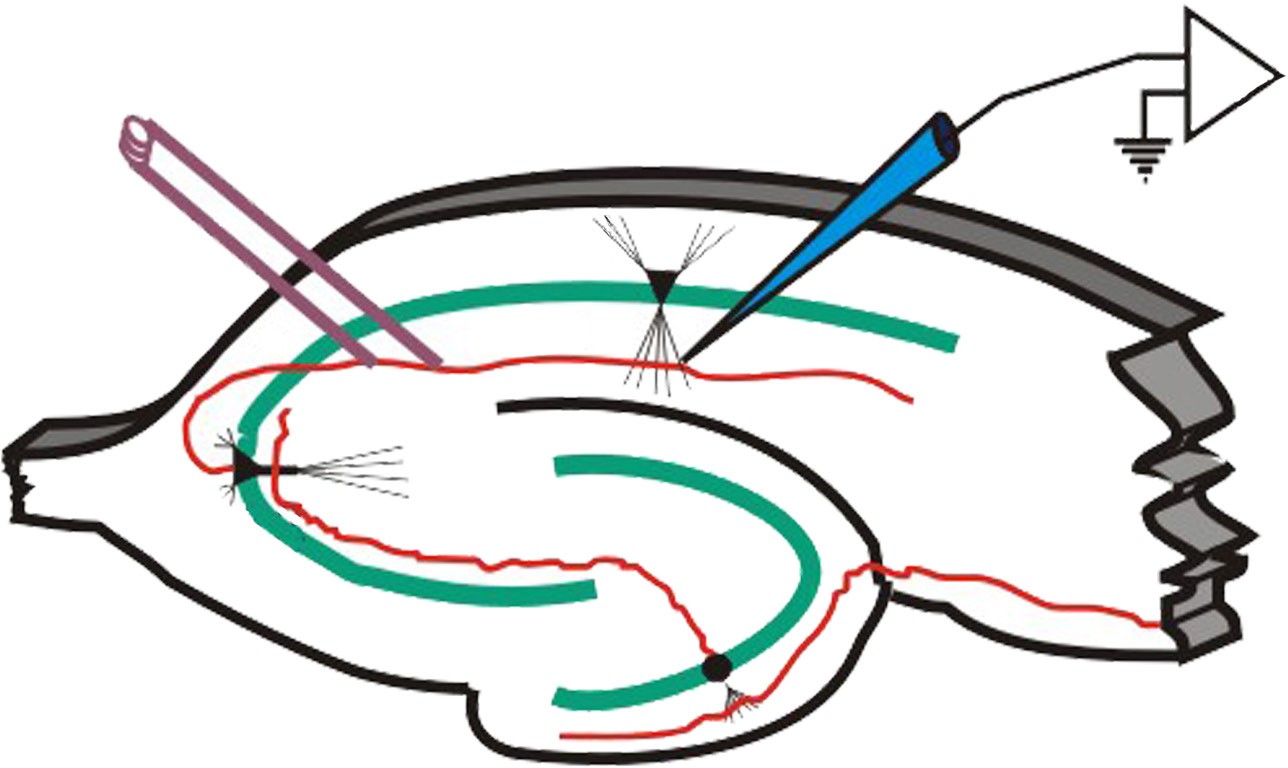
In 1973, Bliss and Lomo were the first to publish evidence of plasticity using electrophysiology. The experiment began at the hippocampal connections of an anesthetized rabbit. They put a stimulating electrode among the axons of the perforant pathway, at the entrance to the hippocampus. A second electrode, capable of detecting electrical charges in brain tissue, was placed among the cells of the dentate gyrus, the area where those axons release neurotransmitter. By stimulating the perforant pathway, Bliss and Lomo could record how neurons of the dentate gyrus respond. A single pulse caused the neurons to depolarize, a measurable observation called a field excitatory post-synaptic potential (fEPSP). The more neurons that depolarize, the larger the fEPSP would be.
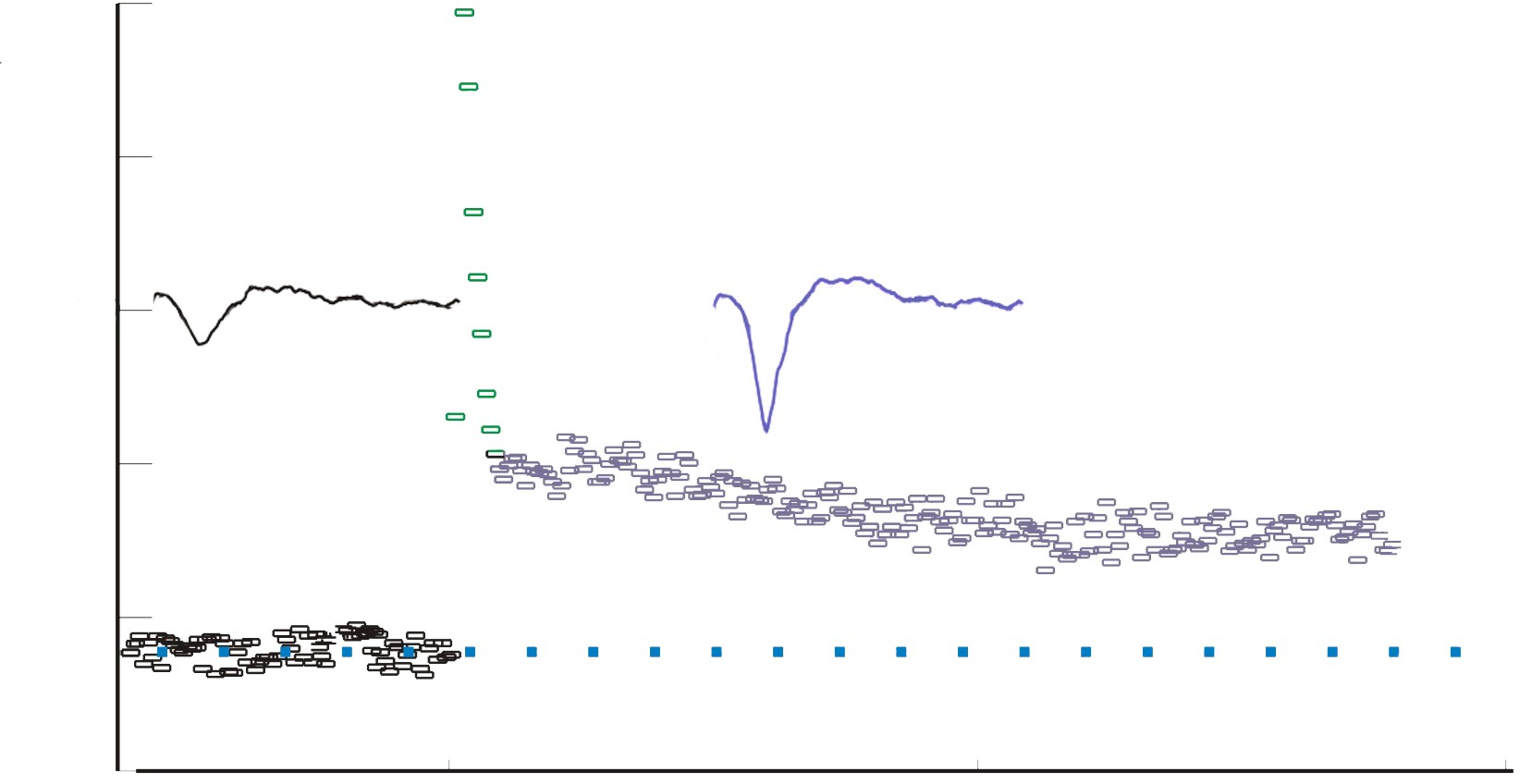
Instead of simply giving a single electrical stimulation, however, Bliss and Lomo were interested in testing Hebb’s theory about plasticity. If “cells that fire together, wire together,” then perhaps they could experimentally drive those cells to fire in a pattern that would induce a rewiring of the connections, resulting in LTP. The duo delivered a very intense electrical stimulation, zapping the axons at 100 stimulations a second (100 Hz) for 3 seconds. This high frequency stimulation (HFS) led to an enhancement of the amplitude of the fEPSP in response to a single stimulus – this demonstrated that LTP was a measurable phenomenon. In a different experimental setup, this LTP was shown to persist up to one year later! In humans, we theorize that some synaptic connections may remain potentiated for our entire lifetime, however investigating this is in humans is ethically constrained.
Long-lasting changes in synaptic strength, such as the LTP that Bliss and Lomo demonstrated, are made possible through a series of molecular and cellular level changes. One form of LTP results from a change in the types of glutamatergic receptors. Of the three classes of ionotropic glutamate receptors, two are important for this form of LTP: the AMPA and the NMDA receptors. The AMPA receptors are the glutamate receptors that we generally imagine as contributing to excitation (more information in section 5.4). When a molecule of glutamate binds to the active site of this receptor, the ligand- gated ion channel changes and allows cations, mostly Na+, to cross the cell membrane, leading to depolarization.
 Stimulator
Stimulator
Recording electrode
High frequency stimulus
EPSP amplitude
Hippocampal slice
 Long term potentiationBaseline
Long term potentiationBaseline
-15
03060
Time (minutes)
Figure 13.18 Schematic of the recording configuration of Bliss and Lomo’s experiments (left) demonstrating that high frequency activation of the Schaffer collaterals while recording the field EPSP in the CA1 region leads to long term potentiation (right).
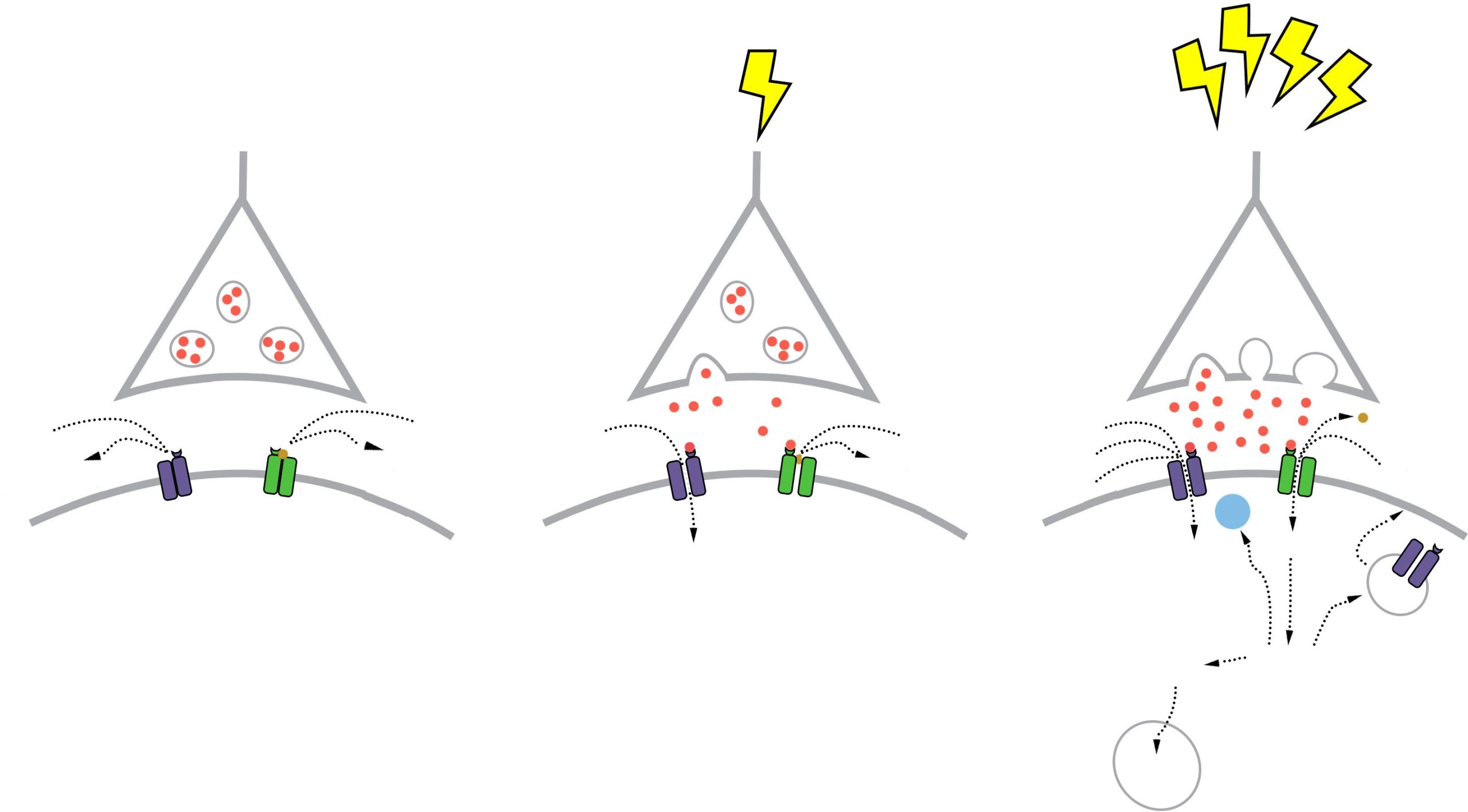
The NMDA receptors are somewhat more complex. NMDA receptors are like the ionotropic AMPA receptors because they are permeable to cations and therefore excitatory, but they have a few specific functional differences. For one, their molecular pore has space for a magnesium ion (Mg2+) to sit in the middle of the ion channel. Mg2+, like the other ions that we have discussed (chapter 4.2), responds similarly to the forces of the electrochemical gradient. Mg2+ is more highly concentrated outside the cell compared to the inside, and it has two positive charges, so these ions are drawn to the interior of the cell. But, the pore of the NMDA receptor is not large enough to allow the bulky Mg2+ ion to actually cross into or out of the cell membrane. Instead, it stays stuck inside the ion channel. Mg2+ physically takes up so much space that it occludes the movement of other ions across the cell membrane, basically blocking passage of ions through the NMDA receptor.
The other relevant feature of the NMDA receptor is that it is permeable to the Ca2+ ion. Increases in intracellular Ca2+ postsynaptically
is the crucial trigger that leads to the cellular expression of LTP. Ca2+ ions activate an enzyme called calcium / calmodulin-dependent protein kinase II (CaMKII). CaMKII itself has many molecular targets. As a kinase, it’s main molecular action is to phosphorylate proteins. When CaMKII becomes activated, it phosphorylates amino acid residues on the AMPA receptor, which enhances their current passing properties, thereby increasing their response with glutamate present. Secondly, CaMKII also contributes to cellular mechanisms which result in increased trafficking of AMPA receptors to the cell surface. Thirdly, CaMKII interacts with the transcription factor cAMP response element-binding protein (CREB), which can then move into the nucleus and instruct the nucleus to synthesize more of the mRNA that leads to increased synthesis of the AMPA receptors. Taken together, increases in intracellular Ca2+ postsynaptically leads to an enhancement of a signal that persists over the time course of hours: The definition of LTP.
But, these NMDA receptors are not activated by glutamate alone. Because of the
 Mg2+Na+Na+,Ca2+Na+,Ca2+PNa+Na+Ca2+CREBCaMKII
Mg2+Na+Na+,Ca2+Na+,Ca2+PNa+Na+Ca2+CREBCaMKII
No stimulationLow stimulationHigh frequency stimulation
Figure 13.19 Molecular mechanisms explaining postsynaptic LTP. At no stimulation, glutamate (pink) does not strongly activate the AMPA receptors (purple; left). A single presynaptic depolarization causes some glutamate to be released, which activates AMPA receptors, causing postsynaptic depolarization (middle). At high frequency stimulation, significant glutamate release activates AMPA receptors, strongly depolarizing the postsynaptic cell, which causes the Mg2+ ion to leave from the NMDA receptor (green). Ca2+ enters through the NMDA receptor, and can trigger long term changes in the molecular components of the neuron (right).
molecular properties of the NMDA receptors, they need two conditions to be fulfilled before these receptors get activated and Ca2+ moves into the cell membrane:
- A ligand like glutamate must activate the receptor. As with other receptors, there is no activation in the absence of an agonist.
- The postsynaptic cell must also be depolarized. When the cell is at positive potentials, the electrical gradient causes the bulky Mg2+ ion that is stuck in the pore to be repelled by the cell’s interior, which then frees the ion channel for
movement of cations across the cell membrane.
Because both conditions must be met before Ca2+ can trigger plasticity through CaMKII activation, the NMDA receptor can be described as a coincidence detector. These properties can help explain why LTP was only observed after Bliss and Lomo activated the hippocampal slices robustly with their high frequency stimulus paradigm. Strong activation depolarized the axonal fibers, which caused a significant amount of glutamate to be released, activating many of the postsynaptic AMPA receptors. This strong
activation caused the postsynaptic neurons to depolarize, which expels the Mg2+ ion out of the NMDA receptor. At this stage, both conditions are fulfilled, and Ca2+ enters into the postsynaptic cell, which activates CaMKII, triggering LTP.
Some glutamatergic connections between neurons contain only NMDA receptors but no AMPA receptors. Because these postsynaptic cells do not depolarize in response to glutamate release, and no current passes through the NMDA receptor due to the Mg2+ block, these synapses do not change their activity even with glutamate release. These synapses are called silent synapses. As we grow, the number of silent synapses decreases, another aspect of brain development.
Long-term depression
Around the same time LTP was being characterized in the rabbit hippocampus, its cellular opposite, long-term depression (LTD), was also being demonstrated in a different experimental preparation. Through the 60s, psychiatrist Eric Kandel and his colleagues worked with the marine mollusk Aplysia californica. With a nervous system of only 20,000 cells, Aplysia is orders of magnitude simpler than the other model organisms used at the time. Additionally, some Aplysia neurons are huge, up to a millimeter in diameter, which took away the need for highly precise equipment.
Aplysia also has a relatively simple anatomy. It breathes using a half-circle of delicate tissue called the gill, which is guarded by the mantle shelf. They also have an organ called the siphon, a small tube that is used for moving water through the animal. Kandel and his colleagues began their exploration of memory by studying the gill-withdrawal reflex, a defensive motor response behavior. When a stimulus,

Figure 13.20 Aplysia californica was the model organism first used to demonstrate the neuronal level changes that underlie the habituation behavior.
such as a hungry predator (or an experimenter’s paintbrush), grazed the siphon, the Aplysia would reflexively withdraw their gill, as if to protect this vital organ by shrinking away from the threat. However, after repeated brush strokes to the siphon, the sea slugs figured out that the stimulus was completely innocuous, and decreased the strength of gill withdrawal. Kandel and team suggested that this change in behavior was a form of learning.
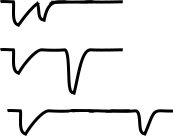
Kandeldiscoveredevidenceofhabituation, the suppression of a normal reflex behavior that is dependent on LTD. To further explore the cellular and molecular level changes behind this LTD, they conducted electrophysiological experiments on Aplysia. The gill-withdrawal reflex circuit relies heavily on two different populations of neurons: the sensory neurons that receive somatosensory information from the skin of the siphon, and the motor neurons that control the muscles of the gill. By using two different tiny glass pipettes, they could impale these neurons, inducing action potential firing in the sensory neuron, and observe changes in membrane potential of the motor
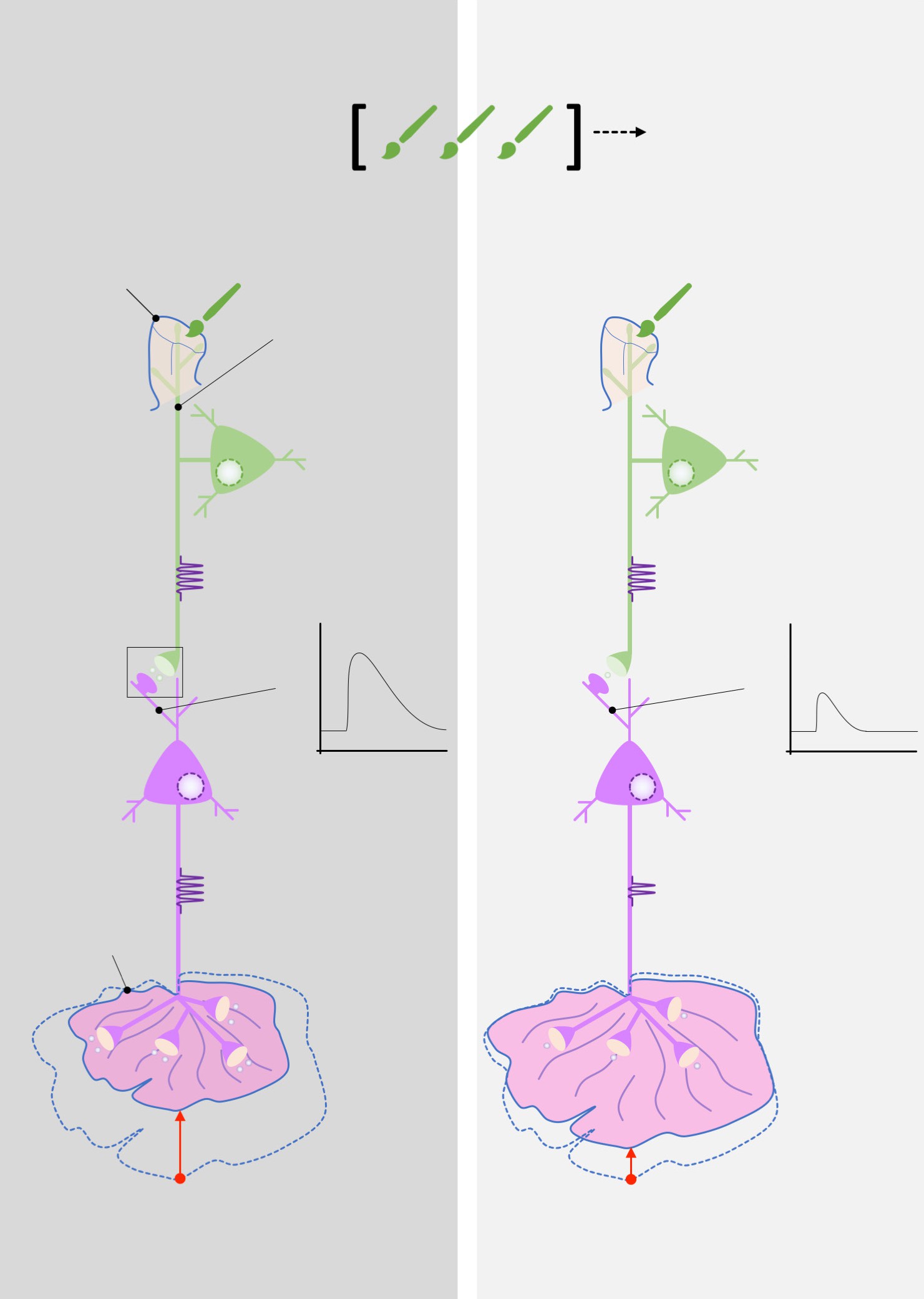 Repeated brush strokesControlHabituatedSiphonSensory neuronLarge EPSPSmall EPSPMotor neuronGillAction potentialsFewer action potentialsStrong withdrawalWeak withdrawal
Repeated brush strokesControlHabituatedSiphonSensory neuronLarge EPSPSmall EPSPMotor neuronGillAction potentialsFewer action potentialsStrong withdrawalWeak withdrawal
neurons (a depolarization of the membrane of a single neuron is called an excitatory post- synaptic potential, or EPSP). When the sensory neuron was activated, they observed an EPSP in response, since an action potential caused release of glutamate that activates post-synaptic receptors on the motor neuron. However, after the reflex had been habituated, the same sensory neuron activation caused a much smaller EPSP in the motor neuron.
The group went about seeing if they could modify this habituated response, curious if a stored memory can be modified by stimuli from the outside world. When they paired the mild siphon touch with a painful electric shock to the tail, the Aplysia began responding with a strong motor reaction, withdrawing the gill very intensely, indicating that the inhibited response disappeared. They called this observation sensitization. In electrophysiological studies, they observed that the EPSP at the motor neuron was much larger following the tail shock.
On a molecular level, presentation of sensitization is downstream of the action of a third population of neurons, interneurons that synapse onto the motor neurons. The noxious stimulus triggers these interneurons
to release the neurotransmitter serotonin, which
Figure 13.21 Habituation of the gill withdrawal reflex was modeled by Kandel using Aplysia. Repeated brush strokes to the siphon caused cellular level changes that led to decreased gill withdrawal.
Oneimportantformofplasticityis
activates the excitatory signaling molecule cAMP found at the terminals of the motor neurons, thus increasing release probability and strengthening the gill withdrawal reflex.
Other forms of plasticity
There are many possible mechanisms that can lead to plasticity. Some activity patterns may induce a short-term plasticity that does not last on the scale of hours or years, but rather on the order of minutes or seconds.
dependent on the activity of the endocannabinoid signaling pathway. During robust post-synaptic depolarization, enzymes such as phospholipase C or DAG lipase create signaling molecules called endocannabinoids (eCBs), which are chemically similar to the primary psychoactive ingredient of Cannabis (chapter 11). These eCBs are neurotransmitters that act in a retrograde manner, meaning they are produced at dendrites and signal to the axons. Expressed at some populations of axons are inhibitory Gi/o coupled
receptors that respond to the presence of eCBs, called cannabinoid receptors (CB1 is the main form that exists in the brain). These receptors inhibit cAMP signaling, which decreases release probability. This specific mechanism of plasticity is called endocannabinoid-mediated LTD.
Another atypical neurotransmitter, the gas nitric oxide (NO), also contributes to a similar form of LTD. The enzyme nNOS synthesizes NO, which diffuses across cell membranes and can activate an intracellular receptor called soluble guanylate cyclase (sGC). SGC changes the activity of the signaling molecule cGMP, which can either lead to enhancement or depression of synaptic strength.
There are also two directionally-opposite forms of plasticity that function at the millisecond timescale, called paired-pulse facilitation (PPF) and paired-pulse depression (PPD). These forms of plasticity can be observed by stimulating a presynaptic area while recording simultaneously from the post-synaptic site. However, instead of delivering a single depolarizing pulse pre- synaptically, these forms of plasticity can only be observed by giving two pulses right after each other, usually separated in time by 10 milliseconds up to a second. In response to these depolarizations, you can also observe two EPSPs. Depending on the time between the pulses (called the interevent interval, or IEI) you may see that the second pulse is a different amplitude than the first. Dividing the amplitude of the second EPSP by the amplitude of the first EPSP gives us a value called the paired pulse ratio (PPR). If the amplitude of the second pulse is increased relative to the first pulse (PPR > 1), this is described as a PPF, whereas if the amplitude in response to the second pulse is smaller than the first (PPR < 1), this is PPD. Both of these plasticity phenomena can be observed
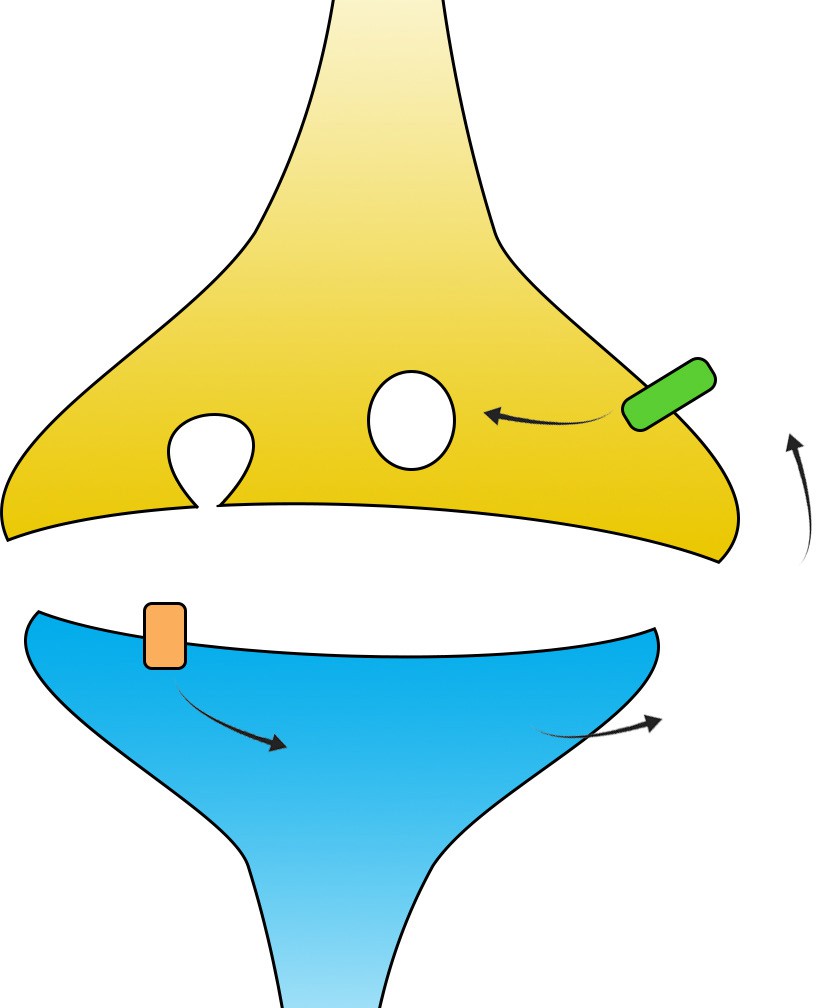 Decreased release probabilityCB1ReCB productionElevated[Ca2+]i
Decreased release probabilityCB1ReCB productionElevated[Ca2+]i
Figure 13.22 Endocannabinoid-mediated LTD is a result of a retrograde signal that decreases release probability.
at the same synapse by varying the duration of
the IEI.
By understanding the molecular mechanisms of synaptic release, we can explain how PPF and PPD can occur at the same synapse. Recall that increased Ca2+ concentration is an important pre-synaptic intracellular signal that allows for vesicular fusion and neurotransmitter release (see section 5.2 for more details). Generally, pre-synaptic Ca2+ enters through voltage-gated calcium channels (VGCCs) that open during depolarization. Delivering a single pulse causes some Ca2+ entry, but giving two pulses, one right after another, allows for more robust activation of the VGCCs thus increasing the intracellular Ca2+ greatly, leading to more neurotransmitter release.
Paired pulse ratio
1.5
1.25
1
.75
20 40
20
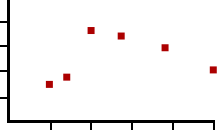
 40
40
100
6080 100
Interevent interval (ms)
Figure 13.22 Paired pulse ratio an electrophysiological measure of short term plasticity.
PPD can also be accounted for by looking at the localization of vesicles in the axon terminal. The readily-releasable pool are the vesicles that are close to the inside of the cell membrane. During a depolarization of the terminal, these vesicles are the first ones to fuse. However, the readily-releasable pool of vesicles can get depleted. On the second pulse, there may be a small number of vesicles remaining because vesicles need to be refilled before they can be released, which makes the amplitude of the second pulse small compared to the first. This is an explanation for paired-pulse depression.
The Open Neuroscience Initiative is funded by a grant from the Vincentian Endowment Fund of DePaul University.
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License.
Cover art: https://pixabay.com/photos/handmade-birthday-cake-dessert-3989069/
- https://en.wikipedia.org/wiki/File:Henry_Gustav_1.jpg
- https://commons.wikimedia.org/wiki/File:Hippolobes.gif https://commons.wikimedia.org/wiki/File:Hippocampus_and_seahorse_cropped.JPG
13.5 https://pixabay.com/vectors/pencil-sharp-school-supplies-153561/
13.6 https://commons.wikimedia.org/wiki/File:Classical_Conditioning_Diagram.png modified by Austin Lim
- https://commons.wikimedia.org/wiki/File:CajalHippocampus.jpeg modified by Austin Lim
- https://commons.wikimedia.org/wiki/File:MorrisWaterMaze.svg modified by Austin Lim https://commons.wikimedia.org/wiki/File:Simple_Radial_Maze.JPG
- https://commons.wikimedia.org/wiki/File:Amygdala.png
- https://commons.wikimedia.org/wiki/File:202003_Model_animal_mouse_mono.svg https://pixabay.com/vectors/sound-audio-music-icon-studio-2935370/
- https://commons.wikimedia.org/wiki/File:Ventral-dorsal_streams.svg modified by Austin Lim
- https://commons.wikimedia.org/wiki/File:Gray727_fusiform_gyrus.png modified by Austin Lim https://commons.wikimedia.org/wiki/File:Fusiform_face_area_face_recognition.jpg https://pixabay.com/photos/man-water-wet-male-face-swimming-984504/ modified by Austin Lim
- https://commons.wikimedia.org/wiki/File:The_structures_of_the_basal_ganglia.png
- https://commons.wikimedia.org/wiki/File:Examples_of_Grid_Cells_with_Different_Grid_Spacing_and_Field_Size.jpg
- https://commons.wikimedia.org/wiki/File:Mark_Hamill_(right)_and_me_(left)_(210269249).jpg https://commons.wikimedia.org/wiki/File:Mark_Hamill_(1980).jpg https://commons.wikimedia.org/wiki/File:Star_Wars_ characters_at_Madame_Tussaud.jpg
https://commons.wikimedia.org/wiki/File:Retrato_del_Maestro_Yoda.jpg https://commons.wikimedia.org/wiki/ File:Leonardo_DiCaprio_2014.jpg https://commons.wikimedia.org/wiki/File:Keanu_Reeves_2014.jpg
- https://commons.wikimedia.org/wiki/File:LTP_exemplar.jpg modified by Austin Lim
- https://commons.wikimedia.org/wiki/File:Potential_mechanisms_of_LTP_in_spinal_dorsal_horn_in_vivo.jpg modified by Austin Lim
- https://commons.wikimedia.org/wiki/File:Aplysia_californica.jpg
- https://commons.wikimedia.org/wiki/File:Habituation.png
- https://commons.wikimedia.org/wiki/File:Alzheimer%27s_disease_brain_comparison.jpg
- https://commons.wikimedia.org/wiki/File:Amyloid-plaque_formation-big.jpg modified by Austin Lim
- https://commons.wikimedia.org/wiki/File:TauProtein.jpg modified by Austin Lim
- https://upload.wikimedia.org/wikipedia/commons/a/a8/Alcohol-induced_brain_damage_%28IA_ alcoholinducedbr00hunt%29.pdf modified by Austin Lim
- https://commons.wikimedia.org/wiki/File:Contrecoup.svg