03-3 Synapse Formation & Elimination
Andrew Bean, PhD
Synapse Formation & Elimination
9.1 Introduction
Perhaps the most remarkable feature of the nervous system is the accuracy of its synaptic connections. The networks of circuits formed by neuronal interactions are responsible for the generation of behavior. Synapse formation is finely regulated. It involves processes at the cellular and subcellular levels, which result in: axons finding their appropriate targets from an array of choices; synapses being formed on the correct cellular compartment; and formation of pre- and postsynaptic specializations that allow for efficient information transfer.
We will discuss the following topics:
- Axon Pathfinding – how axons find their way to specific targets.
- Target recognition – how do axons “know” they have found the correct target?
One general property of neurons is that they migrate from the sites at which they begin to differentiate to their final residence in the nervous system.
- Some neurons migrate to these final positions before they extend processes (e.g., motor neurons).
- Other neurons extend axons as they migrate (e.g., cerebellar granule neurons).
|
Figure 9.1 |
 Granule cells in the developing cerebellum migrate long distances along the processes of radial glial cells; this migration begins in the external granule layer and ends at the granule cell layer. For example, epithelial cells at the luminal surface of the neural tube proliferate to give rise to neuroblasts; some neuroblasts become radial glial cells and extend their processes from the luminal to pial surface. During the development of the cerebellum, granule neurons migrate through the molecular layer along the processes of radial glial cells. A number of neurological mutants have provided invaluable information about neuronal migration. For example, in the “weaver” mutant mouse the genetic defect is in granule neurons and not radial glia. In weaver, the granule cells are not able to migrate to their normal position below the Purkinje cell layer. This defect results in abnormal synaptic circuitry and impaired motor behavior.
Granule cells in the developing cerebellum migrate long distances along the processes of radial glial cells; this migration begins in the external granule layer and ends at the granule cell layer. For example, epithelial cells at the luminal surface of the neural tube proliferate to give rise to neuroblasts; some neuroblasts become radial glial cells and extend their processes from the luminal to pial surface. During the development of the cerebellum, granule neurons migrate through the molecular layer along the processes of radial glial cells. A number of neurological mutants have provided invaluable information about neuronal migration. For example, in the “weaver” mutant mouse the genetic defect is in granule neurons and not radial glia. In weaver, the granule cells are not able to migrate to their normal position below the Purkinje cell layer. This defect results in abnormal synaptic circuitry and impaired motor behavior.
Once neurons have migrated to their final position, and sometimes before, they begin to extend axons. Neurons extend axons because of a specialized structure at the end of axons called growth cones. Granule cells in the developing cerebellum migrate long distances along the processes of radial glial cells. This migration begins in the external granule layer and ends at the granule cell layer.
9.2 Axon Pathfinding
|
Figure 9.2 |
Neurons differentiate from multipotent stem cells and migrate to their final residence in the nervous system. When these neurons reach their residence, they extend an axon and dendrites to send and receive (respectively) information from other neurons. In general, dendrites remain relatively close to the cell body of the neuron, whereas axons may travel long distances to enable interneuronal communication.
Since axons may travel long distances and must search out their target from among many possibilities, the growing end of the axon, or growth cone, must have a mechanism to sense its surroundings.
The distal tip of a growing axon is called the growth cone. Actin is highly concentrated in both the lamellipodia and the finger-like extensions (filopodia). Although the direction of growth cone movement is influenced by the extracellular environment, the growth cone itself possesses intrinsic mechanisms that enable forward movement. Actin is polymerized at the leading edge of the growth cone and moved towards the rear where is it depolymerized. This continuous cycling movement of polymerized actin away from the leading edge towards the rear and the movement of actin monomer to the leading edge again generates a “tank-tread” type of movement. If this actin-based movement is linked to the substrate on which the growth cone is moving, the “tank-tread” movement is endowed with traction and growth cone movement ensues.
|
Figure 9.4 |
Ramón y Cajal first described growth cones and observed that they move in a circuitous route towards their targets. This observation suggested that the growth cones play an active role in the pathfinding process. Roger Sperry later showed that after lesions of the optic nerve in the frog, retinotectal axons regenerate, find their targets, and make synaptic connections with precision. Based on these observations, Sperry suggested the existence of surface markers that are used by the growth cones for pathway and target recognition. Later studies in grasshopper embryo, Drosophila, chick, and zebrafish showed that growth cones follow specific pathways in a variety of species. Since axon pathfinding is similar in disparate species, the mechanisms underlying axon guidance are likely conserved. Understanding the cellular and molecular mechanisms that determine the guidance of growing axons is important because it underlies the initial wiring of the nervous system, but also because it is necessary if there is to be regeneration of function after injury.
9.3 Sources of Guidance Information
|
Figure 9.5 |
Axonal growth cones serve to sense environmental cues and direct the movement of axons in their quest to make synapses with their targets. Guidance cues are necessary to control the growth of axons for long distances to precise destinations. The molecular mechanisms by which these cues act probably involve altering the rate or location of actin polymerization by acting on receptors on the growth cone surface, that are linked to intracellular signaling cascades.
9.4 Guidepost cells
Axonal trajectories appear to be broken up into a series of smaller movements. The axon finds intermediate targets that act as choice points. Axons slow and transform their morphology at these points, and apparently look for another round of directions. Pioneering growth cones (the growth cones on initial axons that are later joined by other axons to create a large axon bundle) are thought to be influenced by these intermediate targets (called guidepost cells in insects). The pioneering growth cones are also influenced by other cues such that the intermediate targets exert only one of a number of influences on the path of growing axons.
9.5 Fasciculation
|
Figure 9.6 |
The bundling together of axons into tracts is called fasciculation. Axonal fasciculation is also a guidance cue as segments of some axonal trajectories migrate along preexisting axon tracts. Thus, axons may follow the pathways laid down by the pioneering axons, although there is some selectivity. An axon may pass a number of axon tracts and make specific choices about which of these axon tracts to follow. It is not yet clear whether the pioneer cells are absolutely required for pathfinding in follower cells. If pioneer cells are not present, the follower cells are more prone to make pathfinding errors, although these errors are largely corrected and the axons find their targets.
9.6 Guidance Forces
- Local vs. long range
- Attraction vs. repulsion
- Target derived signals
Guidepost cells and preexisting axons can affect the outgrowth of developing axons. In addition to these influences, axons also receive long-range guidance information. This short- or long-range guidance information can be attractive or repulsive and serves to push or pull the growth cone in the correct direction towards its target.
Short-range guidance mechanisms, like the interaction with guidepost cells and fasciculation, involve contact-mediated detection by the growth cone filopodia. This contact may be attractive or repulsive. Long- range, diffusable factors may also influence axonal guidance by both attraction and repulsion. It is likely that axons are affected by both local and long-range forces acting simultaneously.
|
Figure 9.7 |
Figure 9.8 |
9.7 Target Recognition-Topographic Maps
Axonal projections are often topographically organized, such that neighboring neurons in the region of origin project to neighboring neurons in the target region (e.g., retinal ganglion cells project topographically onto neurons in the superior colliculus). This organization presents further complexity to the rules of axon outgrowth and raises the question of how the topographic information is encoded. The original experiments of Roger Sperry and later work by many other laboratories has led to the notion that positional information is found at the level of the target region. What is this positional information? One possibility is that each axon has a label that corresponds to an identical label on its appropriate target, allowing specificity of connections. This seems unpractical; there would need to be a very large number of labels, and there is no clear targeting mechanism for the axons. A more likely option was suggested by Sperry in his Chemoaffinity Hypothesis that suggests that positional information is encoded in the form of gradients of signaling molecules at the target which would be detected by complementary gradients of receptors on the axons. This model implies that positional information can be encoded by a small number of molecules and that this information can be “read” by axons as they maneuver in the target region. This chemotropic guidance of axons by diffusable factors that are secreted by cells can involve attractive or repulsive forces.
9.8 Molecules Involved in Axon Growth
Short- and long-range cues act to guide axon growth. These guidance effects can be attractive or repulsive. Axons may use several, or all, of these forces for navigation of single segments of movement. For example, a repellent from behind will “push” an axon into a corridor marked by permissive local cues and bounded by repulsive factors, while an attractant at the end of the corridor may “pull” the axon toward its target. After it was clear that these types of forces work to direct axon growth, attention turned to identifying the molecules that mediate these guidance effects.
9.9 Cell Adhesion Molecules and Extracellular Matrix Molecules
In the past ten years, some of the molecules on the surface of neurons and the substrates on which they grow have been identified. Many of the molecules are responsible for general adhesive interactions between a growth cone and its environment, and others help in choosing which surfaces axons grow on.
Many of the molecules involved in adhesion are glycoproteins and belong to three major families of proteins:
- The immunoglobulin (Ig) superfamily; one major member of this family is the neuronal cell adhesion molecules or N-CAMs (their adhesion properties are Ca2+-independent). There are at least 50 CAMs in mammals.
- The cadherins (their adhesion properties are Ca2+-dependent)
- The integrins; they mediate interactions between the cell-surface and the extracellular matrix (ECM)
Components of the ECM that bind integrins include:
- Fibronectin (secreted by fibroblasts and found in the peripheral nervous system [PNS])
- Laminin (comprises the basement membrane of non-neuronal cells and the basal lamina of cells in the PNS; there appears to be a family of laminins)
Local guidance cues are provided by cell surface proteins that act on the growing axon as it follows its path towards its eventual target.
The homophilic interaction of the CAMs has suggested a role in mediating axon fasciculation and adhesion of axons to cells and/or axon pathways.
|
Figure 9.9 |
Figure 9.10 |
Figure 9.11 |
|
Figure 9.12 |
Figure 9.13 |
Other Short Range Cues:
The ventral midline of the nervous systems acts as an important intermediate target for axons. Growth cones are able to sense specific cues at the midline that influence their decision to cross or not to cross. In addition to being a source of attractants and repellents, the ventral midline is also able to alter an axon’s ability to respond to these cues. For example, after crossing the floor plate, axons that were responsive to the chemoattractant Netrin 1 prior to crossing are, after crossing, no longer able to respond to this cue.
The growth of single sensory axons in an in vitro system led to the identification of an activity in extracts of embryonic spinal cord and of postnatal and adult brain, that promotes the elongation and formation of extensive branches by these axons. Biochemical purification of the activity led to the identification of an amino-terminal fragment of Slit2 as the main active component. Thus, Slit proteins may function as positive regulators of axon collateral formation during the establishment or remodeling of neural circuits. The receptor for slit is present on axonal growth cones and has been identified as the Roundabout (Robo) protein.
|
Figure 9.14 |
9.10 Additional Molecules Involved in Axon Guidance
Receptor tyrosine kinases, extracellular matrix molecules, and netrins, semaphorins and their receptors are also involved in axon guidance. The functions of each are explained below.
9.11 Receptor Tyrosine Kinases
Receptor tyrosine kinases can function to modulate or guide axon growth. In general these receptors are found on developing axons and allow the axon to respond to various growth factors in the local environment. For example, one type of fibroblast growth factor receptor (FGFr, a tyrosine kinase) is necessary for FGF or N-CAM-induced neurite outgrowth. The neurotrophins, NGF, BDNF, NT-3, and NT-4/5, are usually thought of as survival factors for neurons, although these growth factors can also promote axon outgrowth, allow axons to invade their target regions, and promote axon terminal arborization.
9.12 Extracellular Matrix Molecules
Extracellular matrix (ECM) molecules can act to promote or inhibit neurite outgrowth. Laminin, tenascin, collagen, fibronectin, and a number of proteoglycans have been suggested to modulate axonal outgrowth. For example, laminin can promote, while tenascin can inhibit neurite extension. Receptors for ECM molecules include the integrins as well as Ig family members.
The laminin family and its receptors are one of the best studied examples of ECM molecules with regard to neuronal development. There are at least ten different isoforms of laminins that have growth promoting or inhibiting effects on different cell types. The axonal receptors for laminins are integrins. Integrins are heterodimers whose subunit composition determines binding specificity with respect to the laminins. The integrins link ECM signals to the cytoskeleton and various signal transduction pathways. The role of laminins and other ECM molecules in neuronal development is to affect axonal guidance.
|
Figure 9.15 |
9.13 Netrins, Semaphorins and Their Receptors
The netrins are a family of secreted proteins related to laminins in sequence They are both diffusable and cell associated. The netrins were identified using two approaches. A genetic assay in nematode uncovered a mutant gene that resulted in defects in cell migration and axon guidance and an in vitro assay using chick floor plate cell extracts identified a factor that results in outgrowth and guidance of spinal cord axons. Netrins are well conserved from nematode to human and are encoded by only one or two genes in most species. Netrin binds to two types of axonal receptors (DCC/UNC-40 and UNC-5) that determine whether the actions of netrin are attractive or repulsive.
|
Figure 9.16 Axons of primary afferents are directed to termination zones in spinal laminae with the help of a semaphorin III gradient that attracts NT-3 responsive axons (blue) while repelling NGF responsive axons (green), and having no effect on some axons (red) that apparently receive their directional cues from other sources. |
The semaphorins are a large family of cell surface or secreted proteins that mediate growth cone collapse, axonal steering, axonal branching, and axon terminal arborization. At least ten semaphorins are found in humans and they appear to function predominantly as inhibitory (repulsive) factors for axonal guidance. Neuropilins serve as receptors for secreted semaphorins, whereas members of the plexin family are receptors for other semaphorins. Secreted semaphorins do not bind directly to plexins, but plexins can associate with neuropilins. Thus, plexins are receptors for multiple classes of semaphorins, either alone or in combination with neuropilins, and trigger a signal transduction pathway controlling axon guidance.
| Summary of Molecules Involved in Axon Outgrowth and Guidance | |||||
| Ligand | Receptor | Ligand | Receptor | ||
| Contact Attraction | Contact Repulsion | ||||
| Ig CAM | Ig CAM | ||||
| Fasciclin II | Fasciclin II | ||||
| NrCAM | Axonin-1 | Eph Ligand | Eph Receptor | ||
| ECM | Integrin | Semaphorin I | Plexins | ||
| Laminin | Integrin | Slit | Robo | ||
| Chemoattraction | Chemorepulsion | ||||
| Netrins (UNC-6) | DCC (UNC-40, Frazzled) | Netrins (UNC-6) | UNC-5 | ||
| Semaphorins (secreted) | plexins + neuropilins | ||||
Summary
|
Figure 9.17 |
The growth of developing axons depends on:
- Physical guidance cues such as guidepost cells and pioneer axons.
- Local and long-range molecules involved in attraction and repulsion of developing axons (the presence of which substantiates the chemoaffinity hypothesis of Sperry).
- The presence of receptors on developing axons that allow for modulation of growth by environmental factors.
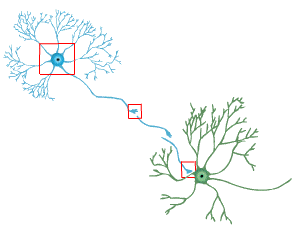
The establishment of synaptic connections and their maintenance depends the interactions of axons and their targets. These interactions are responsible for synapse stabilization and neuron survival. Synaptic transmission depends on the association of the pre- and post-synaptic elements. The pre- and postsynaptic regions of cells are highly specialized architecturally, which allows for efficient information transfer. The architecture of the synapse is formed during development and can change in an activity-dependent manner.
9.14 Molecular Mechanisms of Presynaptic Differentiation
|
Figure 9.18 |
The presynaptic nerve terminal matures from the growth cone and becomes highly specialized to subserve functions necessary for chemical neurotransmission. The presynaptic nerve terminal contains mitochondria and endosomes, although the most striking aspect of this structure is the presence of numerous neurotransmitter-containing synaptic vesicles. Relatively little is known about the molecular mechanisms that underlie differentiation of the presynaptic nerve terminal. It is known that the switch between constitutive vesicle cycling that takes place in the growth cone, and regulated secretion from the mature presynaptic terminal represents a dramatic alteration in function.
9.15 Molecular Mechanisms of Postsynaptic Differentiation
|
Figure 9.19 |
In order for efficient reception of the neurotransmitter signal, the receptors on the postsynaptic cell are clustered at sites opposite the presynaptic active zone. The clustering of postsynaptic receptors is an adaptive behavior that has been studied in detail using the neuromuscular junction as a model system. The nicotinic acetylcholine receptor (AChR) is a ligand-gated ion channel that responds to a signal provided by the neurotransmitter acetylcholine, the transmitter used at the neuromuscular junction. (Additional information about acetylcholine is found in Chapter 11.) AChRs are found at a density of 10,000-20,000/um2 in the postsynaptic membrane while there are about 10/um2 at extrasynaptic sites. The extraordinary concentration of AChRs at the synapse suggests that the muscle has the capacity to anchor these receptors, while their presence in direct opposition to the presynaptic active zone suggests that the nerve plays a role in organizing the postsynaptic membrane.
|
Figure 9.20 |
The cleft separating pre- and postsynaptic membranes contains the basal lamina which coats the entire muscle fiber and is composed of collagens, proteoglycans, and other extracellular matrix molecules including laminin as well as nerve-derived molecules such as agrin (see Chapter 9 part 6). The basal lamina plays a critical role in the assembly of the postsynaptic membrane.
Much of our knowledge about synapse formation comes from studies on nerve-muscle regeneration. Basal lamina patches that are allowed to contact damaged muscle tissue induce the regenerating myotubes to form postsynaptic specializations. These studies suggested that there is an AChR clustering agent in the basal lamina. Agrin was purified from basal lamina extracts as a molecule that is capable of inducing formation of clusters of AChRs on myotubes. Further study showed that agrin is present in motor neurons, is transported down motor neuron axons and is retained in the synaptic basal lamina. Antibodies against agrin inhibit AChR clustering. Agrin deficient mice have disorganized neuromuscular junctions and have decreased number, size, and density of AChRs at the neuromuscular junction. Thus, agrin appears to be a nerve-derived factor that organizes postsynaptic differentiation. Agrin may bind to several candidate receptors including alpha-dystroglycan, heparan sulfate proteoglycans, and a muscle-specific tyrosine kinase (MUSK) to produce its effects.
|
Figure 9.21 |
|
|
Figure 9.22 |
|
|
The synaptic basal lamina possesses proteins that act in pre- and postsynaptic differentiation. A cross section of a normal muscle that contains a nerve terminal directly opposite from the AChRs on the junctional fold of the muscle is shown in Figures 9.21 – 9.22. The basal lamina (BL) ensheaths the myofiber, while the synaptic basal lamina surrounds the synaptic site. The BL serves as a scaffold for regenerating myofibers. In the experiment shown, axons innervating the muscle were damaged and prevented from regenerating and the muscle was injured. New myofibers regenerate within the BL of the original myofiber in the absence of the nerve. In addition, AChRs cluster despite the absence of the nerve. This result shows that the synaptic BL contains signals for clustering AChRs (postsynaptic differentiation). If the motor axon and the muscle are damaged and the muscle is prevented from regenerating, axons regenerate to the original synaptic site in the absence of their targets. Thus, the synaptic BL contains signals for presynaptic differentiation. |
|
The muscle plasma membrane has AChRs on its surface, which are uniformly distributed before the motor nerve growth cone arrives. Shortly after innervation, the distribution of extrajunctional receptors decreases and the density of receptors at the newly forming neuromuscular synapse increases dramatically. This effect occurs precisely at the site where the growth cone releases ACh. These changes occur by the redistribution of existing receptors and the synthesis and insertion of new receptors at synapses.
|
Figure 9.23 |
The axon of the motor neuron induces ACh receptors to cluster on muscle fibers. As the axon approaches the muscle fiber most of the AChRs are distributed diffusely on its surface. When the axon begins to establish synaptic transmission, the distribution of AChRs begins to change. AChRs begin to cluster at the newly formed synaptic contact, while extrasynaptic receptors decrease dramatically.
What factors are responsible for the changes in AChR density during nerve-muscle formation? It is NOT the release of ACh from nerve terminals. Increased AChR density during neuromuscular junction formation results from release of molecules from motor nerve terminals, including AChR inducing activity (ARIA), a molecule that promotes an increase in total AChR and receptor clusters, and Agrin, a protein that induces clustering of pre-existing AChRs and other components of the NMJ.
During nerve-muscle synapse formation, these biological properties of AChRs change after innervation:
- The turnover rate of receptors decreases.
- Their conductance properties increase.
- Receptor open time decreases.
- The subunit composition of individual receptors at the NMJ changes during development.
9.17 Neuronal Survival/Synapse Elimination
At the neuromuscular junction, synapses compete for innervation of a muscle fiber. Although this competition results in a reduction in the number of synaptic inputs to a muscle, the complexity of each synaptic input increases and the strength of each synapse increases.
Some Synapses Are Eliminated During Development
Synapse elimination at the neuromuscular junction involves a loss of AChRs and a withdrawal of the presynaptic terminal. Activity-dependent competition between motor axons favors more active inputs. Direct competition between nerve fibers favors the axon that is able to compete for trophic factors supplied by the target cell. Indirect competition allows the muscle to select the favored axon. An additional strategy for synapse elimination may be that the elimination is provoked by a retrograde messenger. Additional information about retrograde signaling are found in Chapter 14, part 3.
|
Figure 9.24 |
In the adult, almost all muscle fibers are innervated by a single motor neuron. However, early in development many postsynaptic targets are innervated by multiple nerve terminals (this is true for skeletal muscles). During synapse maturation many nerve terminals disappear. Synapse elimination is thought to occur by the withdrawal of presynaptic terminals. Although the number of distinct synaptic inputs to a given target decreases during this process, the complexity of individual remaining terminals actually increases (individual synapses become larger and have more complex structures). The net result of this process is an increase in the overall strength of individual synapses. This process of synapse elimination and rearrangement plays an important role in sculpting the circuitry of the nervous system and the fine tuning of individual synaptic contacts.
The development of connectivity in the central nervous system requires the initiation of synaptic contacts and the maintenance of the correct number of synapses. It is not clear why, but it seems that more synapses are created than are eventually required for proper neuronal function. Under these circumstances, some synapses are eliminated. The process of synapse elimination allows for a “use dependent” form of Darwinism; those synapses that are necessary are retained while those that are simply redundant are removed. The neuromuscular junction and the visual system have been used as models to provide insights into the molecular mechanisms underlying synapse elimination.
|
Synapse elimination can result from activity-dependent signals in the postsynaptic cell. |
||
|
Figure 9.25 |
Figure 9.26 |
Figure 9.27 |
Summary
- A neuron’s ability to reach its target is essential for its survival.
- During development, there is association between the number of presynaptic neurons and the size of their targets.
- Synapse formation is a complex and gradual process.
- Specific molecules from neurons and muscles play important roles in synapse formation and in regeneration (e.g., agrin).
- Finally, synapse elimination and rearrangement result in the fine tuning of neuronal circuitry and synapse strengthening in the nervous system.
9.18 Cell Death, Injury/Regeneration, and Trophic Factors
In most developing tissues, substantial cell death occurs during development. One type of cell death occurs in an activity-dependent manner and is related to synapse elimination (see above). Cell death has been suggested to be the default pathway for all cells, and cells that escape do so by receiving appropriate trophic support. With the growing realization that pathological cell death shares features in common with cell death that occurs during development, a better understanding of developmental cell death may reveal potential therapeutic strategies for the treatment of neurodegenerative disorders and trauma.
The formation of synaptic connections between a presynaptic neuron and its target is often critical to the survival of the presynaptic neuron. In many cases if a synapse is not formed, or if an incorrect synapse is made, then the presynaptic neuron will eventually die. The survival of neurons is often dependent on trophic factors that are produced by the target tissue. Trophic factors are essential for neuron survival; their function is to promote cell growth. (Note: trophic factors should not be confused with chemotropic factors that attract cells.)
9.19 Survival of Certain Neurons Depends on NGF
|
Figure 9.28 |
Early studies on cultured sensory and sympathetic neurons suggested that they require a target-derived factor (nerve growth factor, NGF) for their survival. Competition for NGF occurs when multiple axons innervate a single target that has limited amounts of NGF. Innervating axons possess NGF receptors that bind, internalize, and retrogradely transport NGF. The biochemical cascade produced by NGF internalization and transport results in signals that prevent cell death.
The prototypic trophic factor is NGF; purified NGF is a complex of three protein subunits (alpha, beta, gamma):
alpha: this subunit has an unknown function
beta: this component has biological activity (dimer)
gamma: this subunit has proteolytic activity
One of the first physiological roles that was discovered for NGF was its importance in the survival of sensory and sympathetic neurons. If anti-NGF antibodies are injected into newborn animals to “neutralize” the actions of NGF the survival of sympathetic neurons is greatly reduced.
|
Figure 9.29 |
Figure 9.30 |
|
Figure 9.31 |
For developing neurons in the superior cervical ganglion the natural source of NGF can be blocked by the daily injections of anti-NGF antibodies into newborn animals. This treatment results in the premature death of these neurons. In contrast, if newborn animals are injected with NGF itself, the added NGF actually prevents the naturally occurring death of certain types of neurons.
The cellular actions of growth factors (NGF, BDNF, NT-3, NT-4) are mediated by specific cell surface receptors. The high affinity NGF receptor is a tyrosine kinase that confers the biological activity of NGF to cells by acting through downstream signaling processes that eventually involve alterations in gene transcription. This kinase is known as trkA. Other growth factors bind to different receptor tyrosine kinases. For example, TrkB is specifically activated by low concentrations of BDNF and NT-4/5, and can be activated by high concentrations of NT-3.TrkC is related to trkA, but is most effectively activated by NT-3; all trks are transmembrane proteins that contain an extracellular ligand binding domain and an intracellular (cytosolic) tyrosine kinase domain. When trks are activated by the appropriate ligand (e.g., NGF or BDNF) they initiate a series of intracellular signaling cascades.
|
Figure 9.32 |
|
Figure 9.33 |
NGF is related to other trophic factors such as BDNF, NT-3, NT-4, and CNTF. These factors are structurally related to NGF and are survival factors for neurons that do not respond to NGF. Trophic signals produced by some of these molecules are not all target-derived like NGF. Neurons may receive trophic signals from molecules in the extracellular matrix, from glia, and in an autocrine or paracrine manner. Withdrawal of these factors results in cell death for neuronal populations that are dependent on these factors.
Disruption of different neurotrophin genes in mice has allowed analysis of the role of these factors in sensory-motor pathways. The dorsal root ganglia (DRG) of wild-type mice possess small (red), medium (green), and large (blue) cells. The small cells that carry pain and temperature information are lost when NGF or its high affinity receptor, trkA, are missing. Large cells that innervate muscle spindles are lost when NT-3 or TrkC are missing. These data underline the importance of different neurotrophins in supporting the development of distinct cell populations.
|
Figure 9.34 |
9.20 Neurons in the Peripheral Nervous System Can Regenerate Their Axons
The ability of PNS neurons to regenerate their axons contrasts sharply to damaged neurons in the CNS. This difference is due in large part to the role of trophic factors, which prevent the degeneration of peripheral neurons after axotomy. Recent discoveries have shown that some of the same molecules required for immature neurons to develop and survive are also involved in the survival of adult neurons after injury. What happens when the axons of adult sympathetic neurons are cut?
- If axons of postsynaptic neurons are cut, then presynaptic neurons withdraw their axon terminals.
- If exogenous NGF is added, then regeneration and reinnervation occurs.
- On the other hand, if anti-NGF antibodies are added to neutralize NGF, then regeneration is blocked.
- Moreover, if colchicine (which depolymerizes microtubules) is added to block the retrograde transport of NGF from the target cell to the damaged sympathetic neuron’s cell body, then regeneration is inhibited.
Reactions of sympathetic neurons to injury:
9.21 Neuronal Regeneration
This section focuses on the serious, and often irreversible, damage to central and peripheral nervous systems caused by injury to neurons. In almost every case, neurons have withdrawn from the mitotic cycle and therefore can no longer divide to produce new neurons to replace damaged cells. Therefore, neuron loss due to injury has profound results, since the loss of affected pathways will permanently change the functional circuitry of the nervous system. In those cases where neurons do regenerate following injury, the mechanisms that contribute to regeneration appear to be very similar to the events that promote axon growth and synapse formation during development. Several general rules govern neuronal responses to injury:
- Regeneration in mammals is much poorer than lower vertebrates.
- Regeneration in the CNS is much less likely than in the PNS.
- Trophic factors play important roles in neuronal regeneration.
Neurons in the peripheral nervous system can regenerate their axons. This contrasts with the general inability of damaged neurons in the CNS to regenerate. The regeneration of peripheral neurons is due in large part to the role of trophic factors which provide an important function in preventing their degeneration after axotomy. Some of the same molecules required for immature neurons to develop and survive are also involved in the survival of adult neurons after injury. For example, if the axons of an adult sympathetic neuron are cut, the axon terminals of afferent neurons withdraw their axon terminals. However, if exogenous NGF is added, then regeneration and reinnervation occurs. If anti-NGF antibodies are added to neutralize NGF, then regeneration is blocked. Blocking the retrograde transport of NGF from the target cell to the damaged sympathetic neuron’s cell body also inhibits regeneration. Thus, NGF is required for the regeneration and maintenance of peripheral synaptic connections in the adult PNS.
Schwann cells also contribute to the regeneration of peripheral axons. During damage and regeneration of axons, macrophages are activated and invade damaged areas. They secrete mitogenic factors that stimulate Schwann cell proliferation. Glial cell proliferation results in the increased secretion of extracellular matrix molecules and helps to promote adhesion between the regenerating axons and the remaining supporting cells. The synthesis and secretion of NGF is also stimulated under these conditions and provides important short-term conditions for regeneration. Thus, NGF plays another role in regeneration because it is produced by components of the peripheral nervous system that are not themselves postsynaptic targets.
9.22 Neurons of the CNS Have Very Limited Capacity To Regenerate
Damage to CNS neurons is often irreversible. Why are CNS neurons different than peripheral neurons in this regard? In the adult brain important ECM molecules like laminin and fibronectin are no longer expressed and thus important adhesion molecules are absent from the regenerating environment. In addition, oligodendroglia (the myelinating cells of the CNS) express growth inhibitory molecules in the adult that block axon regrowth.
As discussed above, the environment and properties of peripheral nerves are much more supportive of the regeneration process. For example, Schwann Cells secrete NGF, ECM and other adhesive molecules that are critical for neuronal survival and axon regeneration. In addition, Schwann cells do not express molecules that inhibit axon growth like oligodendroglia in the CNS. The use of peripheral nerve grafts to promote regeneration of damaged nerves in the CNS is one possible way to overcome the limited regeneration potential of CNS neurons.
For example, following transection of the optic nerve, if a section of peripheral nerve is attached to the cut end of the optic nerve and its other end is inserted into superior colliculus, then regeneration of the optic nerve can occur. Evidence supporting the functional regeneration of the cut optic nerves includes:
- Retinal ganglion axons regenerated and reinnervated the superior colliculus.
- New synapses are formed which last for at least one year.
- These new synapses are functional; if retinal neurons are activated by light, then postsynaptic neurons in the colliculus respond (they also respond to direct electrical stimulation of retinal neurons).
- Experimental subjects with PNS nerve grafts have partially restored light avoidance behavior.
- Administration of growth factors during nerve grafts can enhance regeneration.
NGF also appears to play an important role in the survival of cholinergic neurons of the basal forebrain where neurons degenerate in Alzheimer’s disease.
|
Figure 9.40 |
Following damage to an axon by cutting, a sequence of changes occurs during degeneration. Under certain conditions, both retrograde and anterograde transneuronal degeneration can occur.
- Cutting an axon divides it into proximal and distal segments
- Immediately after injury, axons begin to seal off, however, Ca2+ enters damaged axonal elements and activates Ca2+-dependent proteases
- Rapid failure of synaptic transmission occurs within hours (depending on where the axon is cut relative to the distal-proximal axis; metabolic and protein synthetic machinery remains in the cell body)
- Within days presynaptic terminals begin to degenerate and in about 1 week invading glia disrupt and phagocytose synaptic contacts
- The distal segment degenerates slowly and may take up to 1 month; the loss of distal segments is called Wallerian degeneration
- There are also major changes that occur in the neuronal cell body during degeneration; a few days after axotomy, the process of chromatolysis starts in the soma:
- RER (rough endoplasmic reticulum) breaks down and moves to periphery of the cell body (or soma).
- There is an increase in free polyribosomes, mRNA transcription and protein synthesis (the injured neuron’s response to injury is to increase its metabolic rebuilding).
- A damaged neuron’s success in regeneration is linked to its ability to find an appropriate target to reinnervate; failure to contact a target usually results in cell death.
If the injured neuron survives damage (and this often depends on the appropriate trophic factors being present), the process of reinnervation begins:
- The axon’s proximal segment forms sprouts and the process of regrowth begins.
- Axonal sprouts migrate into the conduit formed by the residual endoneural sheath that surrounded the previous axon.
- Axon sprouts migrate to find a target; if a suitable target is found, the neuron survives; if not, the neuron dies.
|
Figure 9.41 |
Cellular damage is not restricted to injured neurons; degeneration is often transneuronal or transsynaptic. In the visual pathway, damage of retinal ganglion neurons often results in degeneration of downstream target neurons in the lateral geniculate and even to neurons in the visual cortex. This is called transsynaptic anterograde degeneration. This process can also happen in the reverse direction (i.e., damage to visual cortex neurons can result in degeneration of geniculate and then retinal neurons), and is called transsynaptic retrograde degeneration.
Traumatic injury to neurons is often irreversible. However, in those instances in which neurons do regenerate following injury, the mechanisms that contribute to regeneration are similar to those events that contribute to axonal growth and synapse formation during development.
During normal development synapse formation is a gradual process that results in one muscle fiber becoming innervated by one motor axon. Following axotomy, the postsynaptic structures called synaptic gutters remain on the surface of the muscle fiber.
During reinnervation, the nerve forms new synaptic contacts at the old synaptic gutters: the one-to-one relationship between nerve and muscle fiber is maintained.
9.23 New Frontiers – Tissue/Cell Transplants
A large number of crippling neurological conditions result from the loss of certain cell populations from the nervous system through disease or injury, and these cells are not intrinsically replaced. Replacement of depleted cell populations by transplantation may be of functional benefit in many such diseases. A diverse range of cell populations is vulnerable, and the loss of specific populations results in circumscribed deficits in different conditions. This diversity presents a considerable challenge if cell replacement therapy is to become widely applicable in the clinical domain, because each condition has specific requirements for the phenotype, developmental stage, and number of cells required.
New discoveries have made inroads in the use of immortalized cell lines to replace or supplement the decreased or lost functions of damaged nervous system tissues. There have also been limited advances in the area of transplantation using fetal tissues (however, this approach is ethically controversial). Areas in which these new approaches have been tried include:
- the synthesis and secretion of catecholamines in patients with Parkinson’s disease
- the synthesis and secretion of growth/trophic factors (e.g., NGF) in patients with Alzheimer’s disease
- the synthesis and secretion of ECM and adhesion molecules to promote regeneration in patients with damaged CNS or PNS tissues
These approaches are technically complicated at both cellular and molecular levels. One of the problems encountered is that although tissue or cell transplants seem to have positive short-term benefits, their long-term effects have been limited.
9.24 Stem Cells
An ideal cell for universal application in cell replacement therapy would possess several key properties: it would be highly proliferative, allowing the ex vivo production of large numbers of cells from minimal donor material; it would also remain immature and phenotypically plastic such that it could differentiate into appropriate neural and glial cell types on, or prior to, transplantation. Critically, both proliferation and differentiation would be controllable. Neural stem cells exist not only in the developing mammalian nervous system but also in the adult nervous system of all mammalian organisms, including humans. Neural stem cells can also be derived from more primitive embryonic stem cells. The cells can be expanded, established in continuous cell lines and differentiated into the three classical neuronal phenotypes (neurons, astrocytes, and oligodendrocytes). The mechanisms that regulate endogenous stem cells are poorly understood. Potential uses of stem cells in repair include transplantation to repair missing cells and the activation of endogenous cells to provide “self-repair. ” Before the full potential of neural stem cells can be realized, we must understand what controls their proliferation, as well as the various pathways of differentiation available to their daughter cells.
9.25 Summary
Postsynaptic targets supply critical neurotrophic factors.
- In addition to NGF, several neurotrophic factors have been identified (e.g., BDNF and neurotrophin 3).
- Reactions of neurons to injury can vary dramatically; neurons usually survive if functional connections are formed during the regeneration process.
- Although regeneration is possible in the PNS, regeneration in the CNS is poor or nonexistent.
Future biomedical research will continue to actively pursue approaches to restore the function of damaged neuronal tissues. The future of understanding important molecules and mechanisms that underlie regeneration lies in basic neurobiological research. Areas to receive attention include:
- The identification of new cell-surface and extracellular matrix molecules that promote axon growth and regeneration.
- The identification and production of new trophic factors that are essential for cell survival and growth.
- The examination of potential use of embryonic stem cells to functionally replace lost neurons.
- In addition, tissue transplantation and peripheral nerve grafts will continue to be studied since they represent one of the most viable approaches to CNS regeneration.
All of the following are guidance cues for axonal outgrowth and pathfinding EXCEPT:
All of the following are guidance cues for axonal outgrowth and pathfinding EXCEPT:
All of the following are guidance cues for axonal outgrowth and pathfinding EXCEPT:
All of the following are guidance cues for axonal outgrowth and pathfinding EXCEPT:
All of the following are guidance cues for axonal outgrowth and pathfinding EXCEPT:
All of the following are guidance cues for axonal outgrowth and pathfinding EXCEPT:
Agrin:
Agrin:
Agrin:
Agrin:
Agrin:
Agrin:
Which of the following is a guidance cue for axonal outgrowth and pathfinding?
Which of the following is a guidance cue for axonal outgrowth and pathfinding?
Which of the following is a guidance cue for axonal outgrowth and pathfinding?
Which of the following is a guidance cue for axonal outgrowth and pathfinding?
Which of the following is a guidance cue for axonal outgrowth and pathfinding?
Which of the following is a guidance cue for axonal outgrowth and pathfinding?
Synapses…
Synapses…
Synapses…
Synapses…
Synapses…
Synapses…
|
Donations to Neuroscience Online will help fund development of new features and content. |

Media Attributions
- Glial Gabby © Google Gemini adapted by Jay C. Brown is licensed under a CC0 (Creative Commons Zero) license