Comparative Cognition
Module 11 Reading
Chapter 10—Comparative Cognition
I. Evolution
A. Evolutionary Theory Before Darwin
B. Basics of Darwinian Evolution
1. Survival of the Fittest
2. Selection
3. Common Descent
4. Punctuated Evolution
5. Criticisms of Evolution
Think Ahead 10-1
6. Moving Beyond Biology: Sociobiology
Check Your Learning 10-1
II. Animal Intelligence
- Historical Study of Animal Intelligence
B. Comparative Cognition
1. Neuroanatomical Differences
2. DNA Markers
Think Ahead 10-2
C. Methodology for Testing Animal Cognition
Check Your Learning 10-2
III. Cognitive Abilities in Nonhuman Animals
A. First, a Warning!
B. Intelligence
1. Assessing Animal Intelligence
2. Nature vs. Nurture
Think Ahead 10-3
C. Theory of Mind
1. Arguments that Animals do Have a Theory of Mind
2. Arguments that Animals do not Have a Theory of Mind
D. Concept Formation
Think Ahead 10-4
1. Object Permanence
2. Conservation
3. Categorization
E. Memory
F. Numerical Skills
Box 10-1 The Natural Selection of Cognitive Skills
1. Telling Time
Think Ahead 10-5
2. Counting and Numerical Abilities
3. Pattern Learning
Check Your Learning 10-3
IV. Cognitive Abilities That Make Humans Unique?
A. Language and Communication
Think Ahead 10-6
B. Tool Use
C. Culture
D. Are Human Cognitive Abilities Truly Unique?
Check Your Learning 10-4
Learning in the Real World: Service Animals
Comparative Cognition
As we’ll learn in this chapter, most scientists agree that all animals are related to each other due to the process of evolution. Because we are related to all other animals (we share common descent), the differences between “us” and “them” is only a matter of degree. To understand the mind of an animal is to understand ourselves. Besides, it’s really fun to talk about what animals can do. Everyone with a dog can tell you just how smart they really can be. Every week some study somewhere reveals some new aspect of animal cognition, its not that animals suddenly got a lot smarter, it’s that scientists are just starting to figure out how to ask the right questions.
As an additional justification for studying animal cognition, a lot of scientists in a lot of fields subscribe to the simple systems approach to science. This means that scientists seek out simpler models first, so that a better understanding of the basic principles can be had before moving onto more complex systems. For psychology, the simple systems approach often includes work with animals, in particular rats, because they are simpler than humans and many more factors can be controlled. I can control what the rat ate for dinner or even if it had any dinner, I can control where it lives, I can control its weight if I want to. All of these factors, which can’t be controlled in humans, contribute to the complexity of understanding behavior and cognition (technically, these factors contribute to error variance, which makes it hard for statistical procedures to detect a true difference; that is, these factors, if left uncontrolled, reduce the power of an experiment). By controlling these factors in animal studies, the scientist has a much easier time recognizing the basic principles of behavior and cognition, presumably the basic principles that all animals share. As a great example of the simple systems approach, Nobel laureate Eric Kandel and his associates have studied classical conditioning in the sea slug (Aplysia californica), a very simple creature, using just three neurons, in order to understand what chemical changes are occurring at the neuronal level during learning (Kandel, 2000).
Insert Picture 1 about here (sea slug)
Evolution
Evolutionary Theory Before Darwin
The Greek philosopher Anaximander was the first to lay out some of the ideas of biological evolution, including notions of common descent and transmutation of species. However, Anaximander also believed that the first humans developed inside of fish (Hergenhahn, 2009)! I guess we had to start somewhere.
As more and more knowledge was building in the fields of biology, geology, and natural history, more and more people began questioning the literal reading of the Book of Genesis which had been a crucial part of the churches stranglehold on scientific thinking throughout the middle ages. Due to overwhelming fossil evidence, there was little question within the scientific community of the 18th century that organisms had changed over time (evolved). The primary question of interest, however, involved the mechanism which drove this change (Gould, 1979). Biologists during the 18th century, led by Jean-Baptiste Lamarck, proposed a theory of evolutionary change in which the individual efforts of an organism within its lifetime could be passed to its offspring (Lamarkian Evolution or inheritance of acquired characteristics). The most famous example involves giraffes who must stretch to reach the high leaves of the Acacia tree (a primary food source for giraffes). Stretching for years will strengthen and lengthen the giraffe’s neck. According to the notion of inheritance of acquired characteristics, these changes to the giraffe’s neck which accrue during its lifetime will be passed along to the giraffe’s offspring (see Figure 10-1). Gould (2002) however, thinks Lamarck got a bad rap in the history books, his critics latched onto a small portion of his thinking and used it as a representation for all of his thinking. Regardless, the notion of the inheritance of acquired characteristics was a very well accepted mechanism to explain evolutionary change, one which was even accepted by Darwin himself for many years (Quammen, 2006). However, we saw this Lamarkian inheritance of acquired characteristics in Chapter 9 when we discussed the possible origins of instincts in social learning so maybe Lamarck’s ideas still retain some relevance.
Insert Figure 10-1 about here
Basics of Darwinian Evolution
Survival of the Fittest
Charles Darwin, during the course of developing his theory of evolution, was inspired by many other scientists. Two of the most notable were Charles Lyell and Thomas Malthus. Charles Lyell wrote The Principles of Geology which convinced Darwin that the world changes with time and that fossils can be dated based on their location in the ground relative to other fossils (stratification). Thomas Malthus wrote An Essay on the Principle of Population in which he described how animals will always produce more offspring than an environment can support. Darwin took from this the notion of the struggle for survival, which later was merged with Herbert Spencer’s ideas to create the notion of “Survival of the Fittest” (Quammen, 2006).
Selection
Modern biologists no longer view the inheritance of acquired characteristics as the mechanism for evolutionary change, but rather natural selection. Natural selection is the process whereby certain characteristics of a species of organisms are “selected” over others. Those characteristics which are selected tend to be those that contribute to the survival of the species, or more specifically, those that increase the probability an animal will mate. Selected characteristics either prolong the life of the individual animal, increase the odds of successful mating, or increase care of resulting offspring. Characteristics which can be selected can be physical traits, social behaviors, or even cognitive abilities.
Insert Figure 10-2 about here
The process of natural selection is driven by environmental pressures. Those traits that increase the likelihood that an organism will survive and reproduce in one environment may not work the same way if the environment changes. The motive behind evolutionary forces is not perfection, but survival in the current environment. This process is demonstrated in Figure 10-2 using the giraffe’s neck length example. When looking at the fossil record we can find millions of different fossil species that are now extinct, and some of these are the same as current species. Changes in the animals over time usually reflect changes in the environment. Humans, during the course of our history have, have been a critical force in the evolution of many animal species. During the history of domestication of a species, humans have used artificial selection (a.k.a. selective breeding) of certain traits; we have become the environment that selects which traits will be passed on the next generation. Wild cows are pretty aggressive animals, however, over thousands of years farmers have selected the trait of calmness in cows so that domesticated cows are now pretty calm. Truth be told though, farmers probably selected against aggression. If it was time to eat a cow, guess which cow the farmer wanted to remove from his or her herd, the most aggressive one (did you ever try to milk an angry cow!). Due to the years of selection for many traits in addition to aggression, modern domestic cows are barely identifiable with their wild ancestors. However, traits that are selected do not have to serve any practical purpose to either the animal or the human breeder, the existence of a multitude of fancy goldfish is a testimony to the fact the artificial selection can be done at the whim of the breeder.
Insert Picture 2 about here (varieties of fancy goldfish)
Common Descent
As stated in the introduction to this chapter, most scientists assume that there are certain physical characteristics, chemical processes, and behavioral processes that are common to all animals. By studying one animal, much can be learned about all animals. This belief is founded on the principle of common descent; if animals can trace back their ancestry to the same point, then they are said to have common descent. In modern biological thinking, all animals share common descent. However, the length of time since a common ancestor was shared is not the same for all animals. For example, humans seem to have a common ancestor with chimpanzees about 6 million years ago, with lemurs about 63 million years ago, and with dogs about 85 million years ago. Since the nearest common ancestor with a chimpanzee is closer in time than the nearest common ancestor with a lemur or dog, humans have more in common with chimpanzees than with lemurs or dogs, while having more in common with lemurs than dogs (Dawkins, 2004). Common ancestry has been a well studied concept for many years. Initially this work was based on physical similarities, but more recently, genetic findings have mostly confirmed earlier beliefs.
Insert Picture 3 about here (Darwin as a monkey-man)
Comparative anatomy is the study of the similarities and differences between different species. If the most recent common ancestor between two species is relatively close in time, the two species have more in common than if the most recent common ancestor is more distant in time. It is fascinating to realize that humans, chimpanzees, lemurs, and dogs all share the exact same organs and bones. In fact, if you look at the bones in the wings of a bird or the fin of a fish, you can find the same bones that are in a human’s hand, though they are shaped differently due to the different functions they perform.
Punctuated Evolution
Gould (1977) describes how Darwin clung to the old belief of Natura non facit saltum which translates to mean “nature does not make leaps”, and made gradualism a critical assumption of his theory. Darwin felt that evolution was an orderly process with a slow and steady progression. Distant ancestors and their descendants could be connected by innumerable transitional organisms. This assumption of Darwinian evolutionary theory almost led to the theory’s demise. The fossil record provides many examples where this assumption does not hold true. Some species disappear from the fossil record very quickly, whereas some species seem to have arisen geologic moment, seemingly out of nowhere. More importantly, the fossil record often failed to provide the crucial intermediate species that Darwin’s beliefs seemed to require. A lack of a “monkey-man” transitional fossil led many millions to criticize evolutionary theory. Darwin argued that change seems abrupt because we are only viewing an imperfect fossil record, one in which most examples are missing, having not yet been discovered or perhaps, due to less than ideal environmental situations at the time that the missing animals should have been becoming fossilized.
Insert Figure 10-3 about here
Gould (1977), however, advocates that gradualism falls short as a description for the evolutionary process and instead argues for punctuated equilibrium (see Figure 10-3). Just as Darwin, Gould argues that evolution is driven by environmental pressures. As long as the environment stays relatively stable, natural selection will ensure that a species fairly quickly evolves those mechanisms most conducive to survival in the current environment. However, environmental changes tend to be fairly rapid, as in the inception of an ice age or the effects caused by the dust stirred up by an asteroid impact. At the time of these rapid changes in the environment, many animals and even whole species, rapidly die out. It is only those organisms that are able to survive and reproduce in the new environment that will pass along their characteristics to the next generation. That is, natural selection will favor a whole new set of characteristics and traits in the new environment. Therefore, the fossil record should consist of long periods of relative stability followed by very short periods of rapid change. Transitional species did exist, but only for a geologic blink of an eye. Because of this, transitional fossils are very rare. Though a definitive “monkey-man” fossil remains elusive, many transitional fossils have definitely been found (Prothero, 2008).
Criticisms of Evolutionary Theory
Though evolutionary theory is widely accepted in the scientific community, it is not without its critics. As already described, a lack of fossils of intermediate species was hard for evolutionary theory to deal with before Gould’s explanation of punctuated equilibrium. A second problem traditionally for evolutionary theory is a lack of experimental evidence for speciation, a problem first described by Romanes (1882). Both supporters and detractors of evolutionary theory accept that a species of animals can be changed radically through the process of selective breeding. Just look at all the different breeds of dogs there are, some are almost unidentifiable as dogs. However, because dogs, regardless of breed, can mate together and have offspring that themselves are fertile, all breeds belong to the same species. Speciation refers to the process whereby a new species is created by evolution. If a new species is developed, then members of the new species will no longer be able to successfully breed with members of the original population.
Think Ahead 10-1
****Because the lifespan of most animals is fairly long and speciation is proposed to be a process stretching across many generations, direct evidence for speciation is elusive. With these limitations in mind, how might a scientist demonstrate speciation in a laboratory?
Recent research has been able to create an artificial speciation in the laboratory using fruit flies (Rice & Hostert, 1993). In one simple demonstration of speciation shown in Figure 10-4, groups of fruit flies were separated and fed either starch based foods or maltose based foods. After keeping the groups separated for eight generations and feeding them these exclusive diets, the resulting flies from the different groups would not mate with each other (Dodd, 1989).
Insert Figure 10-4 about here
Creationism is the belief that all animals, including humans, were created in their current form by a deity. Creationists who are Jewish or Christian often support a literal reading of the book of Genesis. There are many versions of Creationism (e.g. young earth creationism, old earth creationism, etc), some of which are more amenable to scientific evidence than others. Intelligent design is an explanation for scientific evidence that argues that many of the features of the universe came about as the result of an intelligent cause rather than the blind forces of natural selection. Supporters of intelligent design may well believe in many of the basic fundamentals of evolutionary theory, but posit a different mechanism for change.
Moving Beyond Pure Biology: Sociobiology
Traditional evolutionary theory, with its focus of survival and reproduction by the individual, can not explain much of social behavior, in particular altruism which is a concern for the well-being of others which overrides a concern for the self. Sociobiology is an attempt to explain all social behaviors in animals in terms of the evolutionary fitness of these behaviors (Wilson, 1975). Within sociobiology, the primary unit of evolutionary change is not the individual organism, but rather the gene. As Samuel Butler put it so eloquently, “the chicken is the egg’s way of making another egg.” Sociobiology might take this phrase to be, “an organism is a gene’s way of making another gene.”
As more complex social behavior by the organism is added to the gene’s techniques for replicating themselves, altruism becomes increasingly prevalent and eventually appears in exaggerated forms. This brings us to the central theoretical problem of sociobiology: how can altruism, which by definition reduces personal fitness, possibly evolve by natural selection? The answer is kinship: if the genes causing the altruism are shared by two organisms because of common descent, and if the altruistic act by one organism increases the joint contribution of these genes to the next generation, the propensity to altruism will spread through the gene pool. This occurs even though the altruist makes less of a solitary contribution to the gene pool as the price of its altruistic act. (p 3-4)
Though sociobiology is often conceived as a branch of biology, it has much to offer evolutionary psychology. From works such as this comes a study of the evolution of things such as morality, cooperation, and language, truly higher-order social skills that could never be explained using pure evolutionary theory. Evolutionary psychologists view those aspects of humanity which are shared by all such as emotions to be evolved adaptations.
Check Your Learning 10-1
Evolution driven by natural selection provides the basis of modern biology.
Acceptance of evolutionary theory requires the basic assumption of common descent. Animals that share a more recent common ancestor have more in common, both physically and mentally, than animals with a more distant common ancestor.
An understanding of the role of natural selection in adapting to the environment provides a foundation by which much of animal and human behavior can be understood.
Despite the wide acceptance of evolutionary theory, there is a far from universal acceptance of it.
The theory of punctuated equilibrium and the field of sociobiology attempt to answer many of the challenges that evolutionary theory has faced.
Animal Intelligence
Historical Study of Animal Intelligence
In the later part of the 1800’s there was a wealth of interest in the issue of animal intelligence which was a result of Darwin’s later works, The Descent of Man (1871) and The Expression of Emotions in Man and Animals (1872). The field of comparative psychology was founded by George John Romanes (1884; 1889; 1892). Romanes and others felt that there were not only physical similarities between humans and other animals, but cognitive similarities as well. Romanes made some remarkable claims as the nature of animal intelligence and cognition and supported his claims using anecdotal evidence and anthropomorphism. He attributed jealousy and other emotions to fish: “…we are also entitled to attribute to fish the emotions conducive to play; for nothing can well be more expressive of sportive glee than many of their movements. As for jealousy, the fights of many male fish for the possession of females constitutes evidence of emotions which would be called by this name in higher animals” (Romanes, 1892, p. 247). He attributed high levels of intelligence to cats, “Mr. A. Percy Smith informs me of a cat which he possesses, and which, to test her intelligence, he used to punish whenever her kittens misbehaved. Very soon this had the effect of causing the cat herself to train the kittens, for whenever they misbehaved ‘she swore at them and boxed their ears, until she taught the kittens to be clean.’” (p. 414). Stories such as these (and Romanes provided a lot of them) led C. Lloyd Morgan to issue his famous warning known as Morgan’s canon, which we saw in Chapter 9. Essentially, Morgan (1896) warned that Romanes should not assume complicated explanations of animal behavior which require complex cognition when the behavior could be explained using simpler principles.
In the early part of the 20th century, a horse in Germany named Hans became a bit of a celebrity. Hans’ trainer claimed that Hans could perform mathematics, tell time, understand a calendar, read, and spell. Hans would answer questions posed to him by tapping his hoof. If, for example, Hans was asked, “What is 3 X 4?”, he would tap his hoof 12 times (Heyn, 1904). The German government called on psychologist Carl Stumpf to form a panel of experts in various fields to test Hans’ abilities. The entire panel concluded that no tricks were being played and the task of figuring out the basis of Hans’ abilities was passed along to Oskar Pfungst. Pfungst first tried using a questioner other than Hans’ trainer to see if Hans could answer questions even if asked by someone new. Hans passed the tests with flying colors, thus indicating that there was no intentional fraud going on. However, through much further testing, Pfungst found that Hans could only get the right answer if the person asking the question knew the right answer. Could Hans have ESP? No, because not only did the person asking the question have to know the answer, but the questioner needed to be within Hans’ sight in order for him to get the correct answer. Pfungst then decided to examine the behavior of questioners as they were proceeding. Sure enough, as Hans was tapping out the answer, the questioner would unconsciously modify their posture, with a slight shift when Hans had tapped the correct amount. So, instead of being a genius or psychic, Hans, though clever, simply learned to begin tapping his hoof, and stop tapping when he saw the questioner pull back slightly (Pfungst, 1911). Morgan’s canon shines again!
Insert Picture 4 about here (Hans from newspaper)
During a large part of the 20th century, behaviorism was the dominant school of thought in psychology. Behaviorism emerged in large part due to the anecdotal evidence being offered up in the field of comparative psychology in the early part of the 20th century. By definition, behaviorism is concerned with the external behaviors of an organism which can be measured, publicly observable, and replicated. Strict behaviorists such as Skinner, though not necessarily denying the presence of a mental life in animals, felt that the mental life of animals was definitely not something that could be studied experimentally (Hergenhahn, 2009). However, certain behaviorists such as Tolman and Hull proposed a more cognitive approach to learning through the use of intervening variables, mechanisms that are not directly measurable. These intervening variables, which include intelligence, emotions, and motivation, help to explain animal behavior by giving it a cause. For example, a strict behaviorist would say that there is a relationship between hours of food deprivation and amount of food consumed. A cognitive behaviorist would agree with the relationship, but would try to explain it rather than just state it (Zentall, 2002). The hours of food deprivation actually caused hunger (an intervening variable) to increase and it is this increased hunger which caused the amount of food consumed to increase. Modern experimental psychology, with it’s reliance on operational definitions, tends to resemble the cognitive behavioral approach.
Comparative Cognition
Similar to comparative anatomy already discussed is the study of comparative cognition, which is the study of the origins and mechanisms involved in cognition across various species. Just like comparative anatomy, the study of comparative cognition only makes sense with the assumption of common descent. Though the simple systems approach, as described in the introduction, is the approach of choice when studying basic processes felt to be in common with all species (such as the mechanisms of neural conduction), more can be learned about more complex things such as brain anatomy and human cognition and its origins by studying more advanced species such as chimpanzees. Evolutionary thinking has come to dominate the study of comparative cognition with “simpler” and more “primitive” animals having fewer capabilities than “complex” or “advanced” animals (Romanes, 1892; Thorndike, 1911; Shettleworth, 1998).
Neuroanatomical Differences
The brains of humans and other vertebrate animals are similar in a lot of ways. If we were to consider the brain in three basic areas, the hindbrain, the midbrain, and the forebrain, it would only be at the level of the forebrain that we see any major differences and even these seem to be only of differences of degree. The hindbrain and the midbrain together create the brainstem which controls all of those functions needed for basic survival including breathing, heartbeat, arousal, pain, and balance. Even within the forebrain, all vertebrates share similarities in the many areas including the limbic system which is responsible for motivation and emotions. Real differences between humans and other vertebrates only arise when considering the cerebral cortex. The cerebral cortex is the outermost part of the brain that is most responsible for higher order cognitive processes including memory, attention, language, thought, and awareness. Within the cortex there are motor, sensory, auditory, and visual areas. Again, all vertebrates share these areas. As seen in Figure 10-5, where we finally see true differences is in the amount of “gray matter”, those parts of the cortex that do not have a predetermined function and are therefore available for truly higher order thinking (Freberg, 2006).
Insert Figure 10-5 about here
DNA Markers
Recent advances in our understanding of DNA have allowed us to ask questions that were unthinkable a generation ago. When studying the genetic structure of different vertebrates, scientists discovered the human accelerated regions (HARs) which are a set of 49 regions on the chromosome which are very similar across vertebrates, but which are much different in humans.
Think Ahead 10-2
****What biological structure, skills, abilities, and behaviors might be affected by the genes in the HARs? Ask yourself, what are the major things that differentiate humans from other animals?
These highly mutated areas (HARs) are thought to be responsible for the development of human neuroanatomy, language, and complex thought (Pollard et al., 2006). FOXP2 is the name of the gene which is thought to be responsible for the development of language skills. Mutations of the FOXP2 genes in humans have been linked to language development disorders (Vargha-Khadem, Gadian, Copp, & Mishkin, 2005). Mice pups that have been genetically modified to have only one functional FOXP2 gene display significantly less vocalizations than their littermates (Shu et al., 2005).
In addition to the human accelerated regions, since the divergence between humans and chimpanzees (our last common ancestor), many genes have been lost to inactivation in a process called pseudogenization. Genes that used to serve a purpose (and still do in chimpanzees) have simply been turned off, much as a light switch that is turned off has the potential for expression if the switch was only turned on. Genes responsible for smell and the immune response are well represented in these genes that have been inactivated (Wang, Grus, & Zhang, 2006). Other researchers found the genes that code body hair were “turned off” in humans about 240,000 years ago (Winter et al., 2001). Findings such as this allow scientists to ask other questions, including the origins of clothing, which must have gone hand in hand with the loss of body hair. An understanding of the processes of evolution at the level of the gene is necessary in order to even attempt to understand where we came from and who we are.
Methodology for Testing Animal Cognition
Methodology for assessing animal intelligence, as well as other cognitive capabilities, continues to use the techniques which were established in the last century, including mazes and Skinner boxes. Performance measures can be made in mazes and cognitive causes can be inferred. Skinner boxes are ideally suited to measure choice behavior and can be used with almost any species. Skinner boxes can be used to test memory, preferences, or to study a variety of other cognitive skills such as counting or timing (Sturtz, 2008).
Radial-arm mazes are often used to study animal memory. A radial arm maze has a central hub and eight (or more) 4 ft arms that are spaced equidistantly around the hub. A food site is placed at the end of each arm and researchers can then study an animal’s ability to remember which arms it has previously visited (Olton & Samuelson, 1976).
Insert Picture 5 about here (radial arm maze)
A Morris water maze is a tub of murky water which contains an escape platform just below the surface. When a rat is placed in the water, it of course searches randomly at first. However, on subsequent trials the rat’s memory for spatial location can be tested (Morris, 1984).
Hebb and Williams (1946) created an intelligence test for animals with the following disclaimer:
The procedure to be described is certainly not a Binet intelligence test for the rat. All we claim of it is that it differs from a maze-training procedure as a Binet test differs from the learning of nonsense syllables. The method minimizes variations of motivation (either of timidity or eagerness for food); bases its quantitative score on a large number of qualitative analyses of performance; and is economical of time. (p. 59)
In this test (seen in Figure 10-6), a kitchen table was modified such that it had 4 in. high walls around the edges. The entire table was then covered with wire mesh. Additional walls could be added to increase the complexity of the maze though the start and goal boxes always remained in the same locations (Rabinovitch & Rosvold, 1951).
Insert Figure 10-6 about here
Representing the role that biology (in particular ethology) has played in the study of animal cognition, the observation of animals in natural states is also used. A commonly used procedure seen in Figure 10-7, which is most amenable to use with monkeys (though it can be modified for other animals), is called matching to sample. An animal must choose a stimulus which matches a comparison stimulus. With delayed matching to sample, the comparison object is shown, then removed and a delay is added before the animal is allowed to make its selection.
Insert Figure 10-7 about here
However, one of the more interesting techniques for assessing animal intelligence involves setting up problems for animals to solve. If you will recall, the ability to solve problems is one of the key elements of the definition of intelligence. In an interesting example, Weir, Chappell, and Kacelnik (2002) were testing whether crows could successfully choose between straight and bent wires as a tool to be used to remove a bucket containing food from a vertical pipe. One crow removed the bent wire, leaving only the straight wire. A second crow, upon finding only the straight wire, proceeded to take the straight wire and bend it into a hook shape which was then used to retrieve the bucket of food. Another common problem employed by researchers of animal cognition involves the patterned string task which consists of two strings with a food reward clearly attached to the end of one string. The animal must pull the correct string to get the food within their grasp (Harlow & Settlage, 1934). However, some researchers use simpler tools, such as a mirror in the study of the theory of mind.
Check Your Learning 10-2
The study of animal intelligence began in a very nonscientific manner incorporating case studies and anecdotal evidence.
During the majority of the twentieth century the methodology of behaviorism came to dominate the field of animal intelligence.
It is difficult to truly measure those things such as cognitive skills which cannot be directly measured. Psychology needed to latch on to the idea of using operational definitions to make the study of animal intelligence respectable.
When studying cognition in humans, language plays an incredibly valuable role. Unfortunately, animal cognition can only be inferred through changes in behavior.
Our understanding of animal cognition has been limited by the techniques available. New and clever techniques are emerging regularly to help us get a better idea of the limits of animal cognition.
Cognitive Abilities in Nonhuman Animals
First, a Warning!
Throughout the search for cognitive abilities Morgan’s canon must never be forgotten. If an animal’s behavior could be attributable to the presence of a specific cognitive skill or to simpler stimulus-response learning, we must always accept the simplest explanation. Many argue that any “evidence” for animal cognition is the result of wishful thinking on the part of those that see it. Boesch (2007) argues that because of the limitations in methodology and measurement and the huge amount of within species variability (after all, what is the “average” human), any comparison between animal and human cognitive abilities should be avoided due to a bias and only within-species comparisons should be made.
Animal cognition can only be inferred through behavior. Though it is essentially true that human cognition also can only be inferred through behavior, with humans a unique form of behavior is available that is not available in other animals, namely, verbal behavior.
Intelligence
Intelligence is a hypothetical construct that is often used as an explanation for behavior. Originally published as an opinion piece in the Wall Street Journal in 1994, Gottfredson (1997) reports a definition which has been agreed to by 52 intelligence researchers:
A very general mental capability that, among other things, involves the ability to reason, plan, solve problems, think abstractly, comprehend complex ideas, learn quickly and learn from experience. It is not merely book learning, a narrow academic skill, or test-taking smarts. Rather, it reflects a broader and deeper capability for comprehending our surroundings—”catching on”, “making sense” of things, or “figuring out” what to do. (p. 2)
However, more simply, Gottfredson (1998) defines intelligence as the ability to deal with cognitive complexity. It is the second definition that seems more appropriate to define a general characteristic shared by all animals. However, to truly make the definition of intelligence inclusive for all animals, it must include the ability to adapt to changes in the environment.
Assessing Animal Intelligence
As mentioned, intelligence in both humans and animals can only be inferred from behavior. If Johnny completes a spelling test with no mistakes while Billy completes the same test with five mistakes, as long as we know that Johnny and Billy did the same amount of studying (removing experiential factors), we infer that Johnny did better because he is smarter than Billy. Though this inference may be incorrect, it does reflect how the average person thinks. Inferring animal intelligence works in much the same way. A rat could be presented with a “test”, such as a maze, and the rat’s behaviors can be recorded. If rat A completes the maze with fewer errors than rat B, we might infer that rat A has more “intelligence” than rat B.
Nature vs. Nurture
Even though no one really concurs on what intelligence is exactly, a common question of intelligence is, of course, is intelligence a matter of inborn natural abilities (nature), or is intelligence the product of the environment in which one is raised (nurture)?
Think Ahead 10-3
****Do you think that intelligence is the result nature or nurture? Would your opinion on this question be different if we were discussing people versus discussing other animals?
This question is no small issue and is an extension of the earlier empiricism vs. rationalism argument in philosophy. The modern answer is that intelligence is a product of both nature and nurture and the newest questions involve the relative contributions of nature and nurture to intelligence. The question takes on huge social implications when one realizes that there is a correlation between scores on intelligence tests and measures of life success and happiness (Neisser et al., 1996). Are the poor the way they are because they are naturally born with fewer abilities, or are the poor that way because of a lack of opportunities? The argument of nature vs. nurture in intelligence has been played out among animal experiments.
In an attempt to tackle the nature vs. nurture question of intelligence, Heron (1935) selectively bred rats based on their ability to solve a maze. Within only four generations, two strains of rats were produced, maze-bright and maze-dull, which produced significantly different scores on a variety of measures in a maze. Loevinger (1938) reported even more dramatic differences by the 11th generation with no overlap in scores between the group of maze-bright and maze-dull rats. Clearly at least some component of intelligence, as measured in these rats using maze performance, is heritable.
Though Hebb (1949) was probably the first to study the effects of an enriched environment on intelligence. In Hebb’s basic study, shown in Figure 10-8, two groups of rats were reared, one in cages in Hebb’s laboratory (impoverished group) and a second group that was raised in Hebb’s home by his daughters and were allowed to roam the house and play (enriched group). Following several weeks in these conditions, the rats in both groups were tested on standard laboratory tasks including mazes. The rats in the enriched group outperformed the rats in the impoverished group. Mark Rosenzweig and colleagues expanded on this work dramatically (Renner & Rosenzweig, 1987; Rosenzweig, 1984; Rosenzweig, Bennet, & Diamond, 1972). Throughout various studies it was shown that enriched environments (exposure to toys and, depending on the experiment, social interaction) create a multitude of behavioral differences. However, even more interestingly, it was shown that enrichment created a multitude of brain differences compared to impoverished rearing including increased physical weight and volume, increased number of synapses, and increased cerebral blood flow.
Insert Figure 10-8 about here
Brown (1995) attempted to fully tease apart the different aspects of the enriched environments which might be responsible for these behavioral and brain differences. Thirty-day old Long Evans hooded rats (Rattus norvegicus) were exposed to the presence or absence of three factors: object manipulation, social interaction, and observation of other animals manipulating objects. In order to ensure that all rats were as genetically similar as possible, eight male rats who were brothers were mated with eight female rats who were sisters four different times to create the 64 pups used in the experiment. Following 30 days of exposure to these conditions, rats were tested in an open-field (seen in Figure 10-9). An open-field is a square box with lines creating a tic-tac-toe grid. A novel object was placed in the center of each of the squares (except the center square which was the starting point for the rat) and the number of contacts each rat made with these novel objects during a twenty minute period was measured. Contacts with novel objects was felt to be a measure of curiosity, a definite prerequisite for intelligence. It was found that none of these factors (the elements of the enrichment) was necessarily more influential than any of the others. Novel object contacts in the open field as well as amount of locomotion in the open-field were highest for those rats receiving exposure to all three aspects of enrichment and lowest for those rats that received none. Rats receiving two or one of the aspects of enrichment (the exact ones received didn’t seem to matter) scored in between.
Insert Figure 10-9 about here
Theory of Mind
The theory of mind, briefly discussed in Chapter 9 in its relation to imitation, is the ability to attribute thoughts and feelings to oneself and others as well as ability to understand that other’s thoughts and desires are different from one’s own. Clearly this is an ability that is not all or none, it is an ability which can de exhibited in a range. Also, theory of mind is an ability that is subject to development. Young children that display egocentrism (tendency to view the world exclusively from one’s own point of view) probably have not yet fully developed a theory of mind (Piaget & Inhelder, 1947/1968). Among humans, theory of mind is thought to develop alongside language. One of the basic tests for the presence of the theory of mind in children is the ability to recognize that others can have false beliefs about the world (Wimmer & Perner, 1983).
Many question whether the development of the theory of mind in humans requires a genetic endowment or socialization opportunities that are not available to nonhuman animals (Premack & Woodruff, 1978). That is, many question whether nonhuman animals ever develop a theory of mind. Most tests of the theory of mind in humans require the use of words, a child is asked a question and the child gives a response. The response given will then be used to determine if the theory of mind is present. Of course nonhuman animals cannot tell us what is happening, thus making this search particularly challenging.
Arguments that Animals do Have a Theory of Mind
Hare, Call, and Tomasello (2001) demonstrated that subordinate chimpanzees could take advantage of what the dominant chimpanzee knew in order to choose which food containers to approach. The subordinate chimpanzee usually approached the food that was behind a barrier which the dominant chimpanzee could not see. The subordinate chimpanzees needed to consider what the dominant chimpanzee knew and use that knowledge to make an informed decision for itself (see also Hare, Call, Agnetta, and Tomasello, 2000). A similar use of knowledge of others in selecting food sources was demonstrated with dogs (Brauer, Kaminski, Riedel, Call, & Tomasello, 2006)
Hostetter, Russell, Freeman, and Hopkins (2007) showed that chimpanzees appeared to understand that vision plays a central role in attention. Chimpanzees were offered food by human trainers that had either their eyes closed or their eyes open and the chimpanzees clearly produced more visible behaviors when the human trainers eyes were open. Much the same understanding of the role of gazing in human attention was demonstrated with capuchin monkeys, a phylogenetically lower species of monkey (Hattori, Kuroshima, & Fujita, 2007), dolphins (Pack & Herman, 2007), and with dogs (Schwab & Huber, 2006).
Melis, Call, and Tomasello (2006) had chimpanzees play a competitive game with a human opponent. The human player was inside of a box with two different pieces of food which could be retracted by the human player. Each piece of food was accessible from the outside of the box through tubes attached on either side of the box. One tube was clear and the other was opaque. The chimpanzees consistently chose to approach the opaque tube in order to conceal their movements from the human player. In a second experiment, one tube was designed to make noise while the other was silent and the chimpanzees consistently chose the silent one, thus again indicating their attempt to conceal their actions. A similar type of “deception” has been seen with squirrels which use different techniques to bury food when other squirrels are watching compared to when they bury food by themselves (Leaver, Hopewell, Caldwell, & Mallarky, 2007).
Arguments that Animals do not Have a Theory of Mind
Povinelli, Nelson, and Boysen (1990) presented a choice procedure to chimpanzees. There were two experimenters from which food could be requested, one who had seen where the food was hidden and one who was blindfolded and therefore could not know the location of the food. The chimpanzee’s choice of who to request food from was essentially random. Though most studies of theory of mind focus on visual knowledge, human theory of mind incorporates all the senses. Brauer, Call, and Tomasello (2008) found that chimpanzees did not use others’ knowledge of auditory information when making a selection. The chimpanzees were not sensitive to whether or not a competitor could hear where food was being placed, even though they used the information themselves to locate food.
Concept Formation
Concepts are essentially a mental grouping of similar objects, events, or people. Concepts form the foundation of an animal’s cognitive structure, and hence, its understanding of the world.
Think Ahead 10-4
****What benefit do you think an animal might have from the ability to form concepts? Can you envision a world where there were no such thing as a concept?
Concepts allow an animal to successfully interact with their world in many ways; most notably, concepts allow an animal to generalize its learning. If an animal learns that a round, shiny, smooth, and red object tastes good, then the understanding of the concept of “apple” will allow the animal to generalize their knowledge to other apples. Concepts are intimately related to intelligence. The more intelligent an organism, the better they are at forming concepts. A large number of concepts increases an animal’s ability to effectively solve problems (one of the components of the definition of intelligence). The concepts that have been studied the most in humans are conservation and object permanence. However, other more specific concepts, such as “apples” or “fruit” or “food” are also an important part of the cognitive life of an organism.
Insert Picture 6 about here (an apple)
Object Permanence
Object permanence, the recognition that objects that are not longer visible still exist, is an important concept because it allows for a whole set of thinking that is not available without it. New studies are using creative techniques to reveal the origins of object permanence in human infants (Wynn, 1992). Object permanence does not seem to appear suddenly in humans, but instead it seems to develop gradually, with definite evidence for its presence at six months of age. There is evidence for the existence of the concept of object permanence in monkeys (Mathieu, Bouchard, Granger, & Hersovitch, 1976; Collier Baker & Suddendorf, 2006), dogs and cats (Gagnon & Dore, 1994; Dumas & Dore, 1991; see Figure 10-10), and even parrots and jays (Pepperberg & Funk, 1990; Zucca, Milos, & Vallortigara, 2007).
Insert Figure 10-10 about here
Conservation
Conservation, the concept that an object retains its mass or volume despite changes in shape, tends to appear during the toddler years. In Piaget’s classic studies, a child was shown two tall beakers of water which had the same amount of water in them. The child was asked, “Which beaker has more water?” and the kids would answer that they were the same. While the child was watching, the water from one of the beakers would be poured into a shallow bowl. The children were asked a second time, which contains more water, the tall beaker or the shallow bowl. Children that had not yet developed conservation answer that the beaker has more water because it is taller. Though full-fledged conservation doesn’t occur until later (it seems to develop gradually, like object permanence), 3 year-olds tend to succeed on tasks requiring the basic symbolic thinking required for conservation (DeLoache, 1995).
Beran (2007, 2008) found that monkeys (rhesus macaques and capuchin monkeys in this case) could successfully demonstrate conservation of quantity. Monkeys were taught to choose the larger number of objects from among two sets of objects in order to receive a reward. On certain trials, the monkeys were not immediately rewarded, but instead the objects were rearranged such that one or the other sets of objects was made to occupy more space. The monkeys were able to successively choose the larger number of items despite the amount of space they occupied. Conservation was also demonstrated in bonobos, chimpanzees and orangutans (Suda & Call, 2005), and squirrel monkeys (Thomas & Peay, 1976). Conservation has not yet been demonstrated in other animals, though, as has been true in the past with monkeys, this lack of findings could be because other animals do not demonstrate conservation, or because scientists do not yet know how to ask the question.
Categorization
Humans and Animals tend to create categories based on the specific environment that they live in. If an adult human was given a bunch of household objects and asked to sort them into piles, the categories the person would use might be based on the functions of the objects, such as tools in one pile, electrical items in another. Or, the adult might make piles based on their location in the home, such as kitchen items in one pile and bathroom items in another. If a child without as much life experience were asked to perform the same task, they would probably sort using a more “superficial” standard. Perhaps blue objects in one pile and green ones in another or long ones in one pile and square ones in another. Animals, if they could perform this task, would most certainly do it differently still, maybe based on the smell of each item. The categories created would reveal a lot about the way we understand the objects.
Studies have been performed with animals to see if they could understand natural and abstract categories. Bhatt and colleagues showed pigeons pictures of objects belonging to one of four categories (car, cat, chair, or flower). The pigeons were then to press one of four buttons available in a Skinner box, with each button corresponding to a category. The pigeons were able to learn to press the correct buttons when shown the items in the initial set of pictures. However, when shown brand new pictures belonging to one of these four categories, the pigeons were able to choose the correct button better than chance, suggesting that they truly did understand these “natural” categories (Bhatt, Wasserman, Reynolds, & Knauss, 1988). In another study pigeons were shown to be able to successfully group pictures of objects based on human language categories far faster than if the categories were randomly created (Wasserman, Kiedinger, & Bhatt, 1988). Categorization abilities have been displayed in a wide range of animals including such diverse species as honeybees (Benard, Stach, & Giurfa, 2006) and monkeys (Wright & Katz, 2007).
Memory
Memory is the ability to encode, store, and retrieve information, or more to the point for this book, memory is shown whenever evidence of prior learning is demonstrated. In information-processing psychology, the world is seen as being filled with a nearly infinite number of stimuli that could possibly enter the memory system. The memory system, in the form of the sensory register, has access to every single one of these stimuli for a very brief time. However, due to limited a capacity attention, only the most salient of these stimuli ever do enter the memory system. If we pay attention to something, it may move from the sensory register to the short-term memory. The short-term memory is a component of the memory system that holds information for a short time while we use it (a.k.a. working memory). George Miller (1956) reported that short-term memory has a capacity to hold 7 + 2. This figure, though roughly true, has attained near mythical status in psychology. We can take advantage of this when we realize that the magical number seven refers to seven units of information, no matter what the units are. Through the use of chunking information into larger units or other mnemonic strategies, we can greatly increase our memory capacity. Though most of the information in short-term memory is destined to fade away, some of it will get encoded into long-term memory, a nearly limitless capacity for the storage of memories. Eventually, many of these memories will need to be retrieved back to the working memory so we can use them. This process is summarized in Figure 10-11.
Insert Figure 10-11 about here
If we never saw something, then it wouldn’t even be available to the sensory register and we could not “forget” because it never entered the system. However once a stimuli enters the sensory register it is subject to forgetting. If something is in the sensory register, but we fail to pay attention to it, it will not enter short-term memory. If we do pay attention to something and it enters the short-term memory, if we do not actively attend to the information, perhaps through rehearsal, it will fade away. Though the long-term memory seems to have a nearly limitless capacity, information stored can be lost or overwritten. This type of forgetting from long-term memory storage is in some cases a good thing (though I need to remember where I parked my car today at the mall, and I might be able to remember where I parked the last time, I certainly can’t remember where I parked when I went to the mall in 1995!), and in some cases a bad thing (think about patients with Alzheimer’s disease). Later, when we need to retrieve information from the long-term memory to the working memory, we may be unable to access it, as in the case of the tip-of-the-tongue phenomenon.
Studies with animals reveal that animal memory seems to work in essentially the same way as human memory. Although memory processes are similar, the capacity of memory in honeybees and voles (Cooke, Couvillon, & Bitterman, 2007; Ferkin, Combs, delBarco-Trillo, Pierce, & Franklin, 2008) and the duration of memory in cats (Fiset & Dore, 2006) is certainly limited compared to human memory. However, when memory tasks are made more relevant to the animals, as in the case of memory required for foraging or remembering where food is stored, animals perform quite well (Held, Baumgartner, KilBride, & Byrne, 2005; Hoffman & Berna, 2006; Wilkinson, Chan, & Hall, 2007).
Numerical Abilities
In short, animals seem to display many of the same numerical abilities as humans. From an evolutionary perspective, this should not be terribly surprising since all animals face similar challenges that need to be solved, in particular foraging and mating. It is not surprising that similar mechanisms arose to meet similar challenges. However, despite the amazing similarities in animals’ numerical abilities, there are obviously major differences in degree. A rat is never going to solve algebraic equations, no matter how much training you give it, but amazingly, rats do not need to solve algebraic equations to solve any of the challenges they face (see Box 10-1).
Box 10-1The Natural Selection of Cognitive Skills
The environment selects those cognitive skills which allow an animal to solve the problems with which they are faced. Because the majority of species are geographically constrained, the cognitive challenges they face remain relatively stable. However, each animal species is faced with its own unique set of challenges and have therefore evolved unique cognitive skills to meet these challenges. Most cognitive abilities, including counting and timing skills, seem to be most important in foraging situations. An animal that can distinguish more vs. less would know where there is more food and therefore would stand a better chance of survival. Phylogenetically lower animals tend to show fewer numerical skills than more advanced species However, pigeons seem to be able to outshine humans in many ways on mental rotation tasks due to their experiences of viewing the world from many perspectives while flying (Hollard & Dellius, 1982; Kohler, Hoffman, Dehnhardt, & Mauck, 2005). Though a fish might be able to make judgments limited to more vs. less, a chimpanzee might be able to count the objects. Of course humans have taken the counting abilities to the extreme and created fields such as abstract math. As an animal species comes to occupy a larger and more diverse geographical area, as humans have definitely done, they face a larger variety of challenges and therefore need a larger variety of cognitive skills to meet these challenges. The point is that a fish does not need to be able to do algebra in order to find its food and therefore there is no selective pressure for the fish to be able to perform algebra. Animals only evolve those skills that they need to survive.
Telling Time
Think Ahead 10-5
****Obviously, animals have at least some sense of time. They need to know when to eat and sleep. A bee might need to be able to estimate time intervals so that it does not try to gather pollen from the same flower too soon. Obviously, animals can’t use a clock, so what tools do you think animals have at their disposal in order to tell time?
Animals need to be able to discern the time and often use reliable cues such as the sunshine to let them know when a new day (or night) begins. Roosters crow when the sun comes up, but can be fooled to crow in the middle of the night if an appropriate light source is presented. During a solar eclipse, birds stop chirping; presumably they think it is nighttime (Kellogg & Hutchinson, 1964). There is also evidence that many birds use cues from the changing length of day to tell them the time of the year to determine the appropriate time for migration (Helm & Gwinner, 2006). Even humans have reliable circadian rhythms that seem to be reset by the sun in the morning. These are all interesting, but these are circadian timing abilities and their study tends to fall to the domain of biology.
Psychology tends to be more interested in animals’ ability to tell time durations. That is, in the absence of an external cue, can an animal tell time? Not surprisingly, an animals need to tell time in the wild tends to be associated with foraging activities (Bateson, 2003). For example, a bee must be able to remember how long it has been since it visited a flower to assess whether enough pollen has built up to merit a revisit (Skorupski & Chittka, 2006). Plenty of evidence for animals’ ability to tell time exists (see Taylor, 2002 for an interesting example using domestic hens). The simplest come from studies which use Fixed Interval reinforcement schedules primarily using rats and pigeons. If you will recall, during an FI schedule, only a single response in a Skinner box is required by the organism in order to receive reinforcement, however, this response must occur after a fixed amount of time. Whenever animals are placed on this schedule a similar pattern of responding emerges, immediately following reinforcement the animals produce little or no responding. As time passes, the animal’s rate of responding increases, with response rates being maximal just before the reinforcement will be available. Following reinforcement, the pattern repeats. The peak procedure is a technique that takes advantage of this pattern of responding. On some trials no reinforcement is included at the critical time and the animals’ rate of responding is recorded (Roberts & Boisvert, 1998; Batsell, 1991). As the curves in Figure 10-12 show, during an FI 20 s schedule, the animal’s peak rate of responding tends to occur at 20 s, while the peak rate of responding tends to occur at 40 s while in an FI 40 s schedule.
Insert Figure 10-12 about here
After the establishment of the fact that animals can tell time, the next question becomes how do animals tell time? There are two competing theories, the pacemaker and behavioral theories of timing. The pacemaker theory of timing states that animals have an internal clock that pulses at a constant rate and animals tell time by keeping track of this pulsing (Roberts, 1981). The behavioral theory of timing states that animals are able to distinguish intervals of time by remembering and repeating a series of behaviors that fill the interval. For example, if an animal is required to wait at least 5 s before pressing the lever in a Skinner box, it might perform a series of behaviors that it knows were of sufficient length in the past, such as walk to each of the four corners of the box before pressing the lever (Killeen, 1991). However, there is no clear winner among these explanations.
Counting and Numerical Abilities
Counting abilities have been reported in a variety of animal species including primates (Anderson, Stoinski, Bloomsmith, & Maple, 2007; Jordan & Brannon, 2006; Tomonaga, 2008; vanMarle, Aw, McCrink, & Santos, 2006), lemurs (Merritt, MacLean, Jaffe, & Brannon, 2007; Santos, Barnes, and Mahajan, 2005), dogs (Ward & Smuts, 2007), pigeons (Olthof & Santi, 2007), and even salamanders and fish (Uller, Jaeger, Guidry, & Martin, 2003; Agrillo, Dadda, & Bisazza, 2007). The length of this list seems to indicate that numerical abilities were an early arrival in our evolutionary past and serve a crucial function for survival. Of course, the extent of these abilities ranges quite drastically from the mere ability to detect larger vs. smaller in lower animals to the ability to perform theoretical mathematics in humans.
Alex, an African Grey Parrot, was the subject of a thirty-year experiment regarding his numerical and counting abilities. Pepperberg reported that Alex (Avian Learning EXperiment) definitely understood the basic concepts of mathematics at the time of his death in 2007. Alex could distinguish the concepts of bigger, smaller, same, different, over, and under and had even developed an understanding of zero (Pepperberg, 2006a; Pepperberg, 2006b; Pepperberg, 2006c). However, many critics argue that animal numerical abilities are limited at best to a crude understanding of values up to four, far below the levels of numerical skills reported by Pepperberg. Many believe that it is only with language that a true understanding of numbers emerges. So even though Alex did seem to exhibit incredible numerical abilities, these may have been due to his extensive language training. Pepperberg seems to have discovered the numerical skills that African Grey Parrots are capable of, but not what they actually do in the wild.
Pattern Learning
In addition to their counting abilities, animals’ ability to detect and learn patterns would also help them to forage successfully. Hulse and Campbell (1975) repeatedly measured the running time for rats in a maze that consisted of nothing more than a long straight alley. The alley was too long to see the end, but at the end of the alley was a place for reinforcement. Either a 14, 7, 3, 1, or 0 food pellet reinforcement was available at the end of each trial. For one group of rats the size of the reinforcement was chosen randomly on each trial and after several trials, the rats running speed was fairly consistent on each trial. However, in a different group, the size of the reinforcement was systematically changed on each trial, there was a 14 pellet reinforcement on trial 1, 7 pellets on trial 2, 3 pellets on trial 3, 1 pellet on trial 4, then 0 pellets on trial 5. Following these five trials, the pattern repeated 14, 7, 3, 1, and 0. Again, running time was measured on each trial. For rats in the patterned group the running times differed on each trial. On trials when there were 14, 7, or 3 pellets available, the rats ran the alley very quickly, but on trials where 1 or 0 pellets were available, the rats ran the alley very slowly. Apparently, despite being unable to “see” the reinforcement on each trial, the rats in the patterned reinforcement group were able to predict the available reinforcement.
Check Your Learning 10-3
We need to be careful and always remember Morgan’s canon when considering the cognitive capacities of nonhuman animals, animals that do not have language to tell us what they are truly experiencing.
Intelligence, if defined as the capacity to adapt to the challenges of the environment, seems to vary as a direct result of the complexity of the organism.
Evidence of many of the cognitive skills found in humans are also found in animals, particularly higher monkeys such as chimpanzees and gorillas. However, the degree to which these cognitive skills are exhibited seems to depend upon the complexity of the organism and the number of environmental challenges it faces.
When studying animal cognition we should always remember the difference between what degree of cognition they exhibit in the wild and the degree of cognition which they are capable of under carefully controlled laboratory conditions.
Cognitive Abilities that Make Humans Unique?
Early philosophers argued that only humans have a soul, and in this way, God made us special. Then along came Darwin who told us that humans are just another animal, no more special than any other. We share common ancestors with these other animals and, as we’ve seen, we even share many of the same cognitive abilities. What a blow to the collective human ego that was, one whose impact is still being felt. Ever since this revelation, psychologists and anthropologists have been searching for the thing that makes us uniquely human and separates us from the other animals. There are three possible candidates for abilities that may be unique to humans, Language, Tool Use, and Culture. However, to varying degrees, at least some argue that each of these abilities has been demonstrated in animals.
Language and Communication
Communication is the act of conveying ideas and information from one organism to another. All animals communicate in some way or another, dogs wag their tails, cats sometimes hiss, even bees use a waggle dance to let other bees know where they found nectar (von Frisch, 1967). Most forms of animal communication seem to be limited to the expression of information about concrete ideas and events and occur in the present moment. However, many feel that despite these communication abilities, true language is reserved only for humans. Language is a symbolic communication system capable of conveying both concrete and abstract ideas. The use of true language involves the use of grammar, something that seems to be lacking in all animal communication. Language allows the communication of ideas about the past, present, or future as well as the expression of hypothetical possibilities. However, human language can be used in other socially unique ways, as in the transmission of jokes and riddles. It would be hard to imagine a dog telling a joke when its only tools are barking and tail wagging. Additionally, the communication systems of even the most advanced of animals is limited to a few dozen signs whereas human language contains thousands of signs. However, language seems to have evolved from these simpler forms of animal communication. Darwin himself suggested that human language had it roots much earlier in emotional communication (Darwin, 1872).
There is disagreement about when language was first used by humans or their ancestors. Some estimate language emerged about two million (2,000,000) years ago, during the time of Homo habilis, others believe it didn’t emerge until much more recently, forty thousand (40,000) years ago, during the time of Cro-Magnon man. These estimates have been developed from the examination of often incomplete fossilized remains and extrapolating what the larynx would have looked like. Many advances in human speech became possible because of the development of a descended larynx. Very few other animals have evolved a descended larynx. A descended larynx greatly increases the likelihood of choking, therefore, the advantage humans gained by developing speech must have been very large. Many speculate that the advent of speech not only improved communications between humans, but improved social cohesion by creating the ability to plan ahead and work cooperatively (Deacon, 1997; Buss 2008).
Noam Chomsky (1965) proposed that humans are unique among other animals; they have an inborn universal grammar. Inborn universal grammar includes those elements of grammatical structures that are common to all languages. Chomsky believes that these elements of language do not need to be learned, humans are born ready for language through linguistic intuitions. Linguistic intuitions include the recognition that the same sentence can mean different things (i.e. “Visiting relatives can be a pain”), the ability to recognize the parts of speech (i.e. the subject vs. the object), and the ability to recognize when two different sentences mean the same thing (Kellogg, 2003).
In opposition to Chomsky’s assertion that language is a uniquely human skill, many researchers have attempted to teach true language to animals. The earliest attempts involved trying to teach chimpanzees to talk. A chimpanzee named Vicki was raised in a human home along with a regular child. After several years, Vicki acted in many ways like a human child, but not surprisingly her linguistic output was severely limited because she lacked a descended larynx, thus making most human sounds impossible to produce (Kellogg & Kellogg, 1933).
Think Ahead 10-6
****Before reading ahead, how might we ask whether nonhuman animals can use language given that nonhuman animals do not have the appropriate physiology for speech?
Allen and Beatrice Gardner noticed, while watching a film of Vicki, that though she lacked actual words, Vicki did seem to incorporate a lot of gestures in her efforts to communicate with her human “parents”. The Gardners decided that teaching an animal American Sign Language would be a much more sensible alternative.
The Gardners (1989) raised a chimpanzee named Washoe from infancy. Washoe learned much sign language from direct teaching using the principles of reinforcement, but she also learned some of her sign language skills simply by watching the Gardners communicating with each other. Washoe signed during formal training sessions, but also during play times. When Washoe was older, she was given a foster infant named Loulis. In the absence of any reinforcement, Washoe taught Loulis many of the signs she knew. Washoe seemed to view language as something more than a mere tool which could be used to receive reinforcement, language was valuable in and of itself.
Many other monkeys have been taught forms of language as well. Lana, a chimpanzee, was taught an artificial language called Yerkish using plastic symbols (Rumbaugh, 1977). Sarah, a chimpanzee, can manipulate plastic symbols to generate English words. Koko, a lowland gorilla, learned to produce about 1000 words in American sign language and seemed to understand as many as 2000 spoken words. As an interesting side note, Koko had a “nipple fetish” and was accused of sexual harassment with multiple female trainers, charges her lawyer neither accept nor deny. All lawsuits relating to Koko’s alleged harassment have been dropped following an undisclosed settlement in 2005 (No, this is not an April Fools joke or something, Google it and see what you find!).
Despite all these success stories, critics still claim that true language is unique to humans (remember the Fox-P2 gene) because for every success story is a dozen or more failures. Critics argue that though monkeys can be taught language skills, it does not come naturally to them (as opposed to how readily children pick up language with no formal teaching). Also, argue the critics, the monkeys rarely use more than a two “word” phrase and grammatical structure is spotty at best. And finally, critics argue that it in most cases, the monkey’s use of language can be explained as an effort to receive reinforcement (Pinker, 1994).
Tool Use
Natural selection has done a good job giving animals the “tools” they need to solve the problems they face. A great example is Darwin’s famous Galapagos Finches. Long ago finches were isolated on the various islands in the Galapagos Archipelago (32 islands and islets). On each island, the finch’s primary food source is different. On some islands the primary food source is nuts, on other islands it is tree-burrowing insects, on others it is buds and fruits. The forces of natural selection, which work toward solving adaptive problems in the current environment, are different on each island depending on the primary food source. Twelve different, but closely related, species of finches have evolved, each with a different shape of beak (definitely a tool) which is perfectly suited to solve the problems each finch faces. However, the forces of natural selection are relatively slow and sometimes an animal’s natural tools prove insufficient to provide all of the food necessary. In these cases, many animals will supplement their available tools with other objects. It is believed that the ability to use tools has a huge adaptive advantage for animals and that the human thumb evolved to allow us to more efficiently use them.
Insert Picture 7 about here (Darwin’s finches showing beak shapes)
Otters and chimpanzees sometimes use rocks to open nuts. Egyptian vultures use stones to crack open ostrich eggs. Green herons use pieces to food as “bait” when trying to catch fish. Woodpecker finches use pieces of straw to reach into holes and get out tasty insects. However, all of these examples might not qualify as true tool use because the animal has simply used an object it finds. Many researchers argue that true tool use only occurs when an object is modified for a specific purpose (Hauser, 2000).
Jane Goodall’s work has revealed that at least some animals modify objects for a specific purpose. Chimpanzees like to eat termites (don’t we all!). In order to get termites chimpanzees will choose a stick, then strip off the leaves. Further, the chimpanzees will gnaw on the stick to make it more attractive to termites. After preparing the tool, the chimpanzee will then poke the stick into a termite mound and “fish” for termites, occasionally pulling the stick out of the hole in order to suck off any termites that have gotten on the stick (Goodall, 1990). Further, new evidence reveals that the chimpanzees actually seem to use multiple tools in the process. Before the chimpanzee can begin fishing, it first must select a thicker stick which is used to create a decent sized hole in the termite mound through which it can fish (Sanz, Morgan, & Gulick, 2004; Furlong, Boose, & Boysen, 2008). It has also been recently discovered that Chimpanzees will sharpen sticks to use when hunting small mammals (Byrne, 2007). Tool use in lesser monkeys, however, is not as commonly reported (Ducoing & Thierry, 2005).
Insert Picture 8 about here (chimpanzee termite fishing)
Though tool use in nonprimates is much less commonly reported, findings regarding the creation of tools from other animals is beginning to trickle in. As described previously, a New Caledonian crow was able to take a straight piece of wire, insert it into a crack, then use the resulting “hook” to pull a basket containing food from a tube. There have been a lot of other reports, both anecdotal and experimental about crows’ use of tools. Dolphins, generally considered a pretty intelligent animal, have been seen tearing off pieces of sponge which they then wrap around their snout to avoid abrasions when searching for food along the sea floor (Krutzen et al., 2005).
Culture
All around the world, humans are faced with similar problems, but, because historically populations were isolated, these isolated populations solved these problems in slightly different ways. As an example, a similar problem faced humans on both sides of the globe, how to get food to the mouth without getting hands dirty (or food full of germs). The isolated populations in the east and the west solved the problem slightly differently, one created chopsticks, the other forks. Neither solution is better than the other, just different. The accumulation of these different solutions to similar problems creates culture, which, as discussed in Chapter 9, is the byproduct of social learning. Culture refers to ways of doing things and solving problems (involving mores and customs) that are geographically local and passed from one to another within a social group.
Researchers working with Kinji Imanishi, the father of Japanese primatology, studied Japanese snow monkeys extensively during the 1950s. Researchers left sweet potatoes on the beach so they would be able to view the monkeys easily. After doing this for a while, one monkey named Imo, took the sweet potato to the ocean and rinsed off the sand. Soon, this behavior was displayed by other members of the troop. After a few years 80% of the younger monkeys were displaying the ability while only 20% of the older monkeys were (after all, “you can’t teach an old dog new tricks!”). Now, over 50 years later, the tradition has passed on from generation to generation and almost all members of the troop wash sweet potatoes. However, Snow monkeys in different troops on different islands do not show this food washing behavior (de Waal, 2001).
Culture has also been seen in orangutans as evidenced by local expression of distinct behaviors that are passed from generation to generation. The members of at least one group of orangutans makes a sputtering noise with their mouth before going to sleep at night, as though they are saying “good night”. However, this behavior is not seen by orangutans outside this group. Similarly, some groups use leaves as napkins when eating. Orangutans from Sumatra get water from a hole by dipping in a branch, then licking off the leaves while orangutans from Borneo do not exhibit this behavior (Vogel, 2003). Evidence for culture among nonmonkey species has been less prevalent, though evidence for it is mounting (White, Gros-Louis, King, Papakjian, & West, 2007).
Insert Picture 9 about here (an orangutan)
Are Human Cognitive Abilities Truly Unique?
Though humans are clearly “at the top” of the evolutionary ladder, the differences between humans and other animals in every way is merely a matter of degree. Biologically, we share features and functions with other animals. The physiology of humans and other animals seems to have developed out of a shared past which went in different directions depending on the challenges each species faced. Because of these physiological findings, it should not be terribly surprising that human cognition and animal cognition also arose from a shared past. Just like with physiology, each species had its own unique set of challenges to face and the cognitive mechanisms we initially shared with other animal species went in slightly different directions.
Check Your Learning 10-4
Philosophers and psychologists have been searching for many years to find that thing that makes us truly human, especially since an acceptance of evolutionary theory brings with it the conclusion that humans as just another animal. The three most likely candidates in this search for uniqueness are language, tool use, and culture.
Language has been seen as one of the things which truly makes us human, but the distinction between animal communication and human language is not always clear.
Nonhuman primates have been taught many language skills, though many critics dismiss these skills as little more than the product of operant conditioning.
Newer research into the use of tools by animals has revealed some fairly remarkable feats, with animals not only using found objects as tools, but producing them as well by modifying an object.
Evidence of culture is emerging in many different animal species.
The differences between animals and humans seems to be a matter of degree rather than one of kind in every way thus far measured.
Learning in the Real World: Service Animals
Service dogs for people with disabilities (i.e. seeing-eye dogs) have been used for several hundred years at least. Because of the natural intelligence, loyalty, and affection for people, dogs are an obvious choice for people needing assistance. However, a guide dog is not appropriate for all situations. Many other animals are also employed as service animals including “…guide miniature horses, a goat for muscular dystrophy, a parrot for psychosis and any number of animals for anxiety, including cats, ferrets, pigs, at least one iguana and a duck.” The training of most service animals involves the use of the principles of reinforcement to get the animals to perform functions that the disable person is unable to do. However, some of the feats described are quite impressive and seem impossible to train merely using reinforcement, such as a guide pony that leads its blind owner around a street sign which was at just the right height to hit the person in the face. Since the pony would have been able to walk under the sign easily, it needed to demonstrate elements of the theory of mind in order to think about the world from another’s perspective and behave accordingly. However, the most impressive demonstration of animal intelligence for the service of people comes from the use of monkeys (usually capuchin) for people with quadriplegia. Service monkeys can not only perform the specific tasks they have been trained for (and there are a lot of them), but they can modify their learned behavior to fit new situations (Skloot, 2009).
Key Terms
Altruism: A concern for the well-being of others which overrides a concern for the self.
Anthropomorphism: Attribution of human characteristics and tendencies onto nonhuman animals or objects.
Artificial Selection: Intentional breeding of animals to ensure that certain traits or combination of traits which are desirable to the breeder are passed to the next generation.
Behavioral Theory of Timing: Explanation of animals’ ability to tell time that posits that animals tell time by counting the number of behaviors that fit into a particular interval.
Brainstem: Lower two-thirds of the brain, responsible for survival functions including breathing, heartbeat, arousal, pain, and balance.
Category: Class of objects that share something in common; related to concept.
Cerebral Cortex: Outermost part of the brain that is most responsible for higher order cognitive processes including memory, attention, language, thought, and awareness.
Common Descent: If animals can trace back their ancestry to the same point, then they are said to have common descent. In modern biological thinking, all animals share common descent.
Communication: The act of conveying information from one organism to another.
Comparative Anatomy: The study of the physical similarities and differences between different species.
Comparative Cognition: The study of the origins and mechanisms involved in cognition across various species.
Comparative Psychology: The study of the behavior and mental life of nonhuman animals in an attempt to better understand human behavior and mental life.
Concept: An internal representation of something (i.e. object permanence or conservation). Mental grouping of similar objects, events, or people (i.e. categories).
Conservation: The concept that objects retain their mass or volume despite changes in their appearance.
Creationism: Belief that all animals, including humans, were created in their current form by a deity.
Culture: Ways of doing things and solving problems (involving mores and customs) that are geographically local and passed from one to another within a social group.
Evolution: The process of change in a species across generations driven by various forces of biology and nature.
Evolutionary Psychology: Attempt to explain mental and psychological characteristics as the products of natural selection. A direct descendant of the psychological school of Functionalism.
FOXP2 (“forkhead box P2”): A gene thought to be responsible for the human development of grammatical competence and language.
Grammar: A set of rules used for the expression of a language.
Human Accelerated Regions: A set of 49 segments of the human genome that are very similar among all nonhuman vertebrates, but are very different in humans. These segments are thought to be responsible for those things that make us truly human.
Inborn Universal Grammar: Elements of grammatical structure that are common to all human languages
Information Processing Psychology: An approach to cognitive psychology that uses the workings of a computer as a model for understanding the workings of the mind.
Inheritance of Acquired Characteristics: see Lamarkian Evolution
Intelligence: Hypothetical construct often used to explain behavior. Though the exact definition is highly debatable, includes ability to adapt to changes in the environment.
Intelligent Design: An explanation for scientific evidence that argues that many of the features of the universe came about as the result of an intelligent cause rather than the blind forces of natural selection.
Intervening Variables: Mechanisms which are not directly observable, yet which are used to explain the relationship between the independent and the dependent variables in an experiment. Examples in psychology include intelligence and emotion.
Lamarkian Evolution: Notion that an organism can pass along physical changes that have occurred during its lifetime to its offspring (a.k.a. inheritance of acquired characteristics).
Language: A symbolic communication system capable of conveying both concrete and abstract ideas.
Limbic System: Structures in the forebrain that participate in the processes of motivation and emotion.
Long-term Memory: Representations of knowledge and skills which are stored for long periods of time.
Matching to Sample: Procedure for testing animal memory in which a comparison stimulus is presented and the animal must choose the same stimulus in order to receive reinforcement.
Memory: Ability to store, retain, and retrieve information; memory is evident whenever evidence of prior learning has been demonstrated.
Morgan’s Canon: Precept that animal activity should always be explained using the simplest mechanisms possible (a.k.a. principle of parsimony or Occam’s razor).
Natural Selection: Process whereby certain characteristics of a species which lead to the increased probability of successful mating are “selected” over others.
Object Permanence: Concept that objects are permanent entities that do not vanish when out of sight.
Operational Definition: Definition of an intervening variable in terms of the operations used to measure it. As an example, reaction time might be defined as the time between the presentation of a tone and the pressing of a button.
Pacemaker Theory of Timing: Explanation of animals’ ability to tell time that posits an internal “pacemaker” that pulses at a constant rate so that the animal can tell time by keeping track of these pulses.
Peak Procedure: A technique to assess animals timing abilities that withholds reinforcement on certain trials during an FI schedule in a Skinner box and looks at the animals’ response rates.
Punctuated Equilibrium: A theory of evolution that argues that most species remain quite stable for most of their history with the majority of changes occurring in relatively short periods of time driven by radical changes in the environment (new selective pressures).
Radial-Arm Maze: A maze which contains a central hub and eight or more equidistantly spaced arms, each about four feet long. Used primarily to test animal memory.
Rehearsal: The repetition of information in working memory for the purpose of either keeping it for current use, or to ease the transition to long-term memory.
Sensory Register: A component of the memory system that registers everything available to the senses.
Short-term Memory: A set of processes for holding information in our awareness (a.k.a. working memory).
Simple Systems Approach: The approach to science which seeks out the simplest system possible first in order to understand the basic mechanisms which drive all systems, both the simple ones and the complex ones.
Sociobiology: Attempt to explain all social behaviors in animals in terms of the evolutionary fitness of these behaviors.
Speciation: The process whereby a new species is created by evolution.
Survival of the Fittest: Phrase referring to the notion that organisms tend to have more offspring than the environment can support and that there is a competition for limited resources, thus ensuring that those best suited to the current environment have the greatest chances of surviving.
Theory of Mind: Ability to attribute thoughts and feelings to oneself and others as well as ability to understand that other’s thoughts and desires are different from one’s own.
Transmutation of Species: In evolutionary theory, the alteration of one species into another.
References
Agrillo, C., Dadda, M., & Bisazza, A. (2007). Quantity discrimination in female mosquitofish. Animal Cognition, 10(1), 63-70.
Anderson, U. S., Stoinski, T. S., Bloomsmith, M. A., & Maple, T. L. (2007). Relative numerousness judgment and summation in young, middle-aged, and older adult orangutans (Pongo pygmaeus abelii and Pongo pygmaeus pygmaeus). Journal of Comparative Psychology, 121(1), 1-11.
Bateson, M. (2003). Interval timing and optimal foraging. In W. H. Meck (Ed.), Functional and Neural Mechanisms of Interval Timing, (pp. 113-141). Boca Raton, FL: CRC Press.
Batsell, W. R., Jr. (1991). Timing like a rat: A classroom demonstration of the internal clock. Teaching of Psychology, 18(4), 229-231.
Benard, J., Stach, S., & Giurfa, M. (2006). Categorization of visual stimuli in the honeybee Apis mellifera. Animal Cognition, 9(4), 257-270.
Beran, M. J. (2007). Rhesus monkeys (Macaca mulatta) succeed on a computerized test designed to assess conservation of discrete quantity. Animal Cognition, 10(1), 37-45.
Beran, M. J. (2008). Capuchin monkeys (Cebus apella) succeed in a test of quantity conservation. Animal Cognition, 11(1), 109-116.
Bhatt, R. S., Wasserman, E. A., Reynolds, W. F., Jr., & Knauss, K. S. (1988). Conceptual behavior in pigeons: Categorization of both familiar and novel examples from four classes of natural and artificial stimuli. Journal of Experimental Psychology: Animal Behavior Processes, 14(3), 219-234.
Boesch, C. (2007). What makes us human (Homo sapiens)? The challenge of cognitive cross-species comparison. Journal of Comparative Psychology, 121(3), 227-240.
Brauer, J., Call, J., & Tomasello, M. (2008). Chimpanzees do not take into account what others can hear in a competitive situation. Animal Cognition, 11(1), 175-178.
Brauer, J., Kaminski, J., Riedel, J., Call, J., & Tomasello, M. (2006). Making inferences about the location of hidden food: Social dog, causal ape. Journal of Comparative Psychology, 120(1), 38-47.
Brown, J. C. (1995). Environmental stimulation, social rearing, and conspecific modeling affects the behavior of rats. Unpublished master’s thesis, University of Wisconsin, Oshkosh.
Buss, D. M. (2008). Evolutionary Psychology: The New Science of the Mind (3rd ed.). Boston: Allyn and Bacon.
Chomsky, N. (1965). Aspects of the Theory of Syntax. Cambridge, MA: MIT Press.
Collier-Baker, E. & Suddendorf, T. (2006). Do chimpanzees (Pan troglodytes) and 2-year-old children (Homo sapiens) understand double invisible displacement? Journal of Comparative Psychology, 120(2), 89-97.
Cooke, M. H. Y, Couvillon, P. A., & Bitterman, M. E. (2007). Delayed symbolic matching in honeybees (Apis mellifera). Journal of Comparative Psychology, 121(1), 106-108.
Cooper, R. M. & Zubec, J. P. (1958). Effects of enriched and restricted early environments on the learning ability of bright and dull rats. Canadian Journal of Psychology, 12(3), 159-164.
Darwin, C. (1859). On the Origin of Species by Means of Natural Selection. London: Murray.
Darwin, C. (1871). The Descent of Man and Selection in Relation to Sex. London: Murray.
Darwin, C. (1872). The Expression of Emotions in Man and Animals. London: Murray.
Dawkins, R. (2004). The Ancestor’s Tale: A Pilgrimage to the Dawn of Life. Boston: Houghton-Mifflin.
Deacon, T. (1997). The Symbolic Species: The Coevolution of Language and the Human Brain. Hammondsworth, England: Allen Lane.
DeLoache, J. S. (1995). Early understanding and use of symbols: The model model. Current Directions in Psychological Science, 4, 109-113.
de Waal, F. (2001). Without walls. New Scientist, 172(2321), 46.
Dodd, D. M. (1989). Reproductive isolation as a consequence of adaptive divergence in Drosophila pseudoobscura. Evolution, 43(6), 1308-1311.
Ducoing, A. M. & Thierry, B. (2005). Tool-use learning in Tonkean macaques (Macaca tonkeana). Animal Cognition, 8(2), 103-113.
Dumas, C. & Dore, F. Y. (1991). Cognitive development in kittens (Felis catus): An observational study of object permanence and sensorimotor intelligence. Journal of Comparative Psychology, 105(4), 357-365.
Ferkin, M. H., Combs, A., delBarco-Trillo, J., Pierce, A. A., & Franklin, S. (2008). Meadow voles, Microtus pennsylvanicus, have a capacity to recall the “what”, “where”, and “when” of a single past event. Animal Cognition, 11(1), 147-159.
Fiset, S. & Dore, F. Y. (2006). Duration of cats’ (Felis catus) working memory for disappearing objects. Animal Cognition, 9(1), 62-70.
Freberg, L. A. (2006). Discovering Biological Psychology. Boston: Houghton-Mifflin.
Furlong, E. E., Boose, K. J., & Boysen, S. T. (2008). Raking it in: The impact of enculturation on chimpanzee tool use. Animal Cognition, 11(1), 83-97.
Gagnon, S. & Dore, Y. (1994). Cross-sectional study of object permanence in domestic puppies (Canis familiaris). Journal of Comparative Psychology, 108(3), 220-232.
Gardner, R. A. & Gardner, B. T. (1989). A cross-fostering laboratory. In R. A. Gardener, B. T. Gardner, & T. Van Cantfort (Eds.), Teaching Sign Language to Chimpanzees. Albany: State University of New York Press.
Goodall, J. (1990). Through a Window: 30 Years Observing the Gombe Chimpanzees. Boston: Houghton-Mifflin.
Gottfredson, L. S. (1994, December 13). Mainstream science on intelligence. Wall Street Journal, A18.
Gottfredson, L. S. (1997). Forward to “Intelligence and Social Policy”. Intelligence, 24(1), 1-12.
Gottfredson, L. S. (1998). The general intelligence factor. Scientific American Presents, 9(4), 24-29.
Gould, S. J. (1977, May). Evolution’s erratic pace. Natural History, 86(5), 12-16.
Gould, S. J. (1979, Oct). Shades of Lamarck. Natural History, 88(8), 22-28.
Gould, S. J. (2002). The Structure of Evolutionary Theory. Boston: Harvard University Press.
Hare, B., Call, J., Agnetta, B., & Tomasello, M. (2000). Chimpanzees know what conspecifics see and do not see. Animal Behaviour, 59(4), 771-785.
Hare, B., Call, J., & Tomasello, M. (2001). Do chimpanzees know what conspecifics know? Animal Behaviour, 61(1), 139-151.
Harlow, H. F. & Settlage, P. H. (1934). Comparative behaviour of primates VII capacity of monkeys to solve patterned string tasks. Journal of Comparative Psychology, 18(3), 423-434.
Hattori, Y., Kuroshima, H., & Fujita, K. (2007). I know you are not looking at me: Capuchin monkeys’ (Cebus apella) sensitivity to human attentional states. Animal Cognition, 10(2), 141-148.
Hauser, M. (2000). Wild Minds: What Animals Really Think. New York: Henry Holt.
Hebb, D. O. (1949). The Organization of Behavior. New York: Wiley.
Hebb, D. O. & Williams, K. (1946). A method of rating animal intelligence. Journal of General Psychology, 34, 59-65.
Held, S., Baumgartner, J., KilBride, A., & Byrne, R. W. (2005). Foraging behavior in domestic pigs (Sus scrofa): Remembering and prioritizing food sites of different value. Animal Cognition, 8(2), 114-121.
Helm, B. & Gwinner, E. (2006). Migratory restlessness in an equatorial nonmigratory bird. PloS Biology, 4(4), 611-614.
Hergenhahn, B. R. (2009). An Introduction to the History of Psychology. Belmont, CA: Wadsworth.
Heron, W. T. (1935). The inheritance of maze learning ability in rats. Journal of Comparative Psychology, 19(1), 77-89.
Heyn, E. T. (1904, September 4). Berlin’s wonderful horse. New York Times, SM1.
Hoffman, M. L. & Berna, M. J. (2006). Chimpanzees (Pan troglodytes) remember the location of a hidden food item after altering their orientation to a spatial array. Journal of Comparative Psychology, 120(4), 389-393.
Hollard, V. D. & Delius, J. D. (1982). Rotational invariance in visual pattern recognition by pigeons and humans. Science, 218, 804–806.
Hostetter, A. B., Russell, J. L., Freeman, H., & Hopkins, W. D. (2007). Now you see me, now you don’t: Evidence that chimpanzees understand the role of the eyes in attention. Animal Cognition, 10(1), 55-62.
Hulse, S. H. & Campbell, C. E. (1975). “Thinking ahead” in rat discrimination learning. Animal Learning and Behavior, 3, 305-311.
Jordan, K. E. & Brannon, E. M. (2006). Weber’s law influences numerical representations in rhesus macaques (Macaca mulatta). Animal Cognition, 9(3), 159-172.
Kandel, E. R. (2000). Cellular mechanisms of learning and the biological basis of individuality. In E. R. Kandel, J. H. Schwartz, & T. M. Jessell (Eds.), Essentials of Neural Science and Behavior (pp. 161-178). Norwalk, CT: Appleton & Lange.
Kellogg, R. T. (2003). Cognitive Psychology (2nd ed.). London: Sage.
Kellogg, P. P. & Hutchinson, C. M. (1964). The solar eclipse and bird song. Living Bird, 3, 185-193.
Kellogg, W. N. & Kellogg, L. A. (1933). The Ape and the Child: A Study of Environmental Influence Upon Early Behavior. New York: McGraw-Hill.
Killeen, P. R. (1991). Behavior’s time. The Psychology of Learning and Motivation, 27, 295-334.
Kohler, C., Hoffman, K. P., Dehnhardt, G., & Mauck, B. (2005). Mental rotation and rotational invariance in the Rhesus monkey (Macaca mulatta). Brain, Behavior, and Evolution, 66, 158-166.
Krutzen, M., Mann, J., Heithaus, M. R., Connor, R. C., Bejder, L., & Sherwin, W. B. (2005). Cultural transmission of tool use in bottlenose dolphins. Proceedings of the National Academy of Sciences of the United States of America, 102(25), 8939-8943.
Leaver, L. A., Hopewell, L., Caldwell, C., & Mallarky, L. (2007). Audience effects of food caching in grey squirrels (Sciurus carolinensis): Evidence for pilferage avoidance strategies. Animal Cognition, 10(1), 23-27.
Loevinger, J. (1938). “Reasoning” in maze-bright and maze-dull rats. Journal of Comparative Psychology, 26(3), 413-426.
Mathieu, M., Bouchard, M. A., Granger, L., & Herscovitch, J. (1976). Piagetian object permanence in Cebus capucinus, Lagothrica flavicauda, and Pan troglodytes. Animal Behaviour, 24, 585–588.
Melis, A. P., Call, J., & Tomasello, M. (2006). Chimpanzees (Pan troglodytes) conceal visual and auditory information from others. Journal of Comparative Psychology, 120(2), 154-162.
Merritt, D., MacLean, E. L., Jaffe, S., & Brannon, E. M. (2007). A comparative analysis of serial ordering in ring-tailed lemurs (Lemur catta). Journal of Comparative Psychology, 121(4), 363-371.
Miller, G. A. (1956). The magical number seven, plus or minus two: Some limits on our capacity for processing information. Psychological Review, 63(2), 81-97.
Morgan, C. L. (1896). An Introduction to Comparative Psychology. London: Scott.
Morris, R. (1984). Developments of a water-maze procedure for studying spatial learning in the rat. Journal of Neuroscience Methods, 11(1), 47-60.
Neisser, U., Boodoo, G., Bouchard, T. J., Jr., Boykin, A. W., Brody, N. Ceci, S., et al. (1996). Intelligence: Knowns and unknowns. American Psychologist, 51(2), 77-101.
Olthof, A. & Santi, A. (2007). Pigeons (Columba livia) associate time intervals with symbols in a touch screen task: Evidence for ordinality but not summation. Journal of Comparative Psychology, 121(1), 82-94.
Olton, D. S. & Samuelson, R. J. (1976). Remembrance of places passed: Spatial memory in rats. Journal of Experimental Psychology: Animal Behavior Processes, 2(2), 97-116.
Pack, A. A. & Herman, L. M. (2007). The dolphin’s (Tursiops truncates) understanding of human gazing and pointing: Knowing What and Where. Journal of Comparative Psychology, 121(1), 34-45.
Pepperberg, I. M. (2006a). Grey parrot (Psittacus erithacus) numerical abilities: Addition and further experiments on a zero-like concept. Journal of Comparative Psychology, 120(1), 1-11.
Pepperberg, I. M. (2006b). Ordinality and inferential abilities of a grey parrot (Psittacus erithacus). Journal of Comparative Psychology, 120(3), 205-216.
Pepperberg, I. M. (2006c). Grey parrot numerical competence: A review. Animal Cognition, 9(4), 377-391.
Pepperberg, I. M. & Funk, M. S. (1990). Object permanence in four species of psittacine birds: An African grey parrot (Psittacus eritacus), an Illiger mini macaw (Ara maracana), a parkeet (Melopsittacus undulatus) and a cockatiel (Nymphicus hollandicus). Animal Learning and Behavior, 18, 97-108.
Pfungst, O. (1911). Clever Hans (The Horse of Mr. von Osten): A Contribution to Experimental Animal and Human Psychology (Trans. C. L. Rahn). New York: Henry Holt. (Originally published in German, 1907).
Piaget, J., & Inhelder, B. (1948/1967). The Child’s Conception of Space. New York: W.W. Norton.
Pinker, S. (1994). The Language Instinct. New York: William Morrow.
Pollard, K. S., Salama, S. R., Lambert, N., Lambot, M, Coppens, S., Pedersen, J. S., et al. (2006). An RNA gene expressed during cortical development evolved rapidly in humans. Nature, 443(7108), 167-172.
Prothero, D. (2008). What missing link? New Scientist, 197(2645), 35-41.
Quammen, D. (2006). The Reluctant Mr. Darwin. New York: Norton.
Premack, D. G. & Woodruff, G. (1978). Does the chimpanzee have a theory of mind? Behavioral and Brain Sciences, 1, 515-526.
Rabinovitch, M. S. & Rosvold, H. E. (1951). A closed field-intelligence test for rats. Canadian Journal of Psychology, 5(3), 122-128.
Renner, M. J. & Rosenzweig, M. R. (1987). Enriched and Impoverished Environments: Effects on Brain and Behavior. New York: Springer-Verlag.
Rice, W. R. & Hostert, E. E. (1993). Laboratory experiments on speciation: What have we learned in 40 years? Evolution, 47(6), 1637-1653.
Roberts, S. (1981). Isolation of an internal clock. Journal of Experimental Psychology: Animal Behavior Processes, 7(3), 242-268.
Roberts, W. A. & Boisvert, M. J. (1998). Using the peak procedure to measure timing and counting processes in pigeons. Journal of Experimental Psychology: Animal Behavior Processes, 24(4), 416-430.
Romanes, G. J. (1882). The Scientific Evidences of Organic Evolution. London: MacMillan.
Romanes, G. J. (1884). Mental Evolution in Animals. New York: Appleton.
Romanes, G. J. (1889). Mental Evolution in Man. New York: Appleton.
Romanes, G. J. (1892). Animal Intelligence. London: New York: Appleton.
Rosenzweig, M. R. (1984). Experience, memory, and the brain. American Psychologist, 39, 365-376.
Rosenzweig, M. R., Bennett, E. L., & Diamond, M. C. (1972). Brain changes in response to experience. Scientific American, 226(2), 22-29.
Rumbaugh, D. M. ed. (1977). Language Learning by a Chimpanzee: The Lana Project. New York: Academic Press.
Santos, L. R., Barnes, J. L., & Mahajan, N. (2005). Expectations about numerical events in four lemur species (Eulemur fulvus, Eulemur mongoz, Lemur catta and Varecia rubra). Animal Cognition, 8(4), 253-262.
Sanz, C., Morgan, D., & Gulick, S. (2004). New insights into chimpanzees, tools, and termites from the Congo basin. American Naturalist, 164(5), 567-581.
Schwab, C. & Huber, L. (2006). Obey or not obey? Dogs (Canis familiaris) behave differently in response to attentional states of their owners. Journal of Comparative Psychology, 120(3), 169-175.
Shettleworth, S. J. (1998). Cognition, Evolution, and Behavior. New York: Oxford University Press.
Shu, W., Cho, J. Y., Jiang, Y., Zhang, M., Weisz, D., Elder G. A., et al. (2005). Altered ultrasonic vocalization in mice with a disruption in the Foxp2 gene. Proceedings of the National Academy of Sciences of the United States of America, 102(27), 9643-9648.
Skloot, R. (Jan 4, 2009). Creature comforts. The New York Times Magazine, 34.
Skorupski, P & Chittka, L. (2006). Animal cognition: An insect’s sense of time? Current Biology, 16(19), R851-R853.
Sturtz, B. R., Bodily, K., Hernandez, M., Schmidtke, K., & Katz, J. S. (2008). Animal cognition. In S. F. Davis & W. Buskist (Eds.), 21st Century Psychology: A Reference Handbook (pp 285-293). Los Angeles: Sage.
Suda, C. & Call, J. (2005). Piagetian conservation of discrete quantities in bonobos (Pan paniscus), chimpanzees (Pan troglodytes), and orangutans (Pongo pygmaeus). Animal Cognition, 8(4), 220-235.
Taylor, P. (2002). Perception of time duration by domestic hens. Applied Animal Behavior Science, 76(1), 41.
Thomas, R. K. & Peay, L. (1976). Length judgments by squirrel monkeys: Evidence for conservation? Developmental Psychology, 12(4), 349-352.
Thorndike, E. L. (1911). Animal Intelligence. New York: MacMillan.
Tomonaga, M. (2008). Relative numerosity discrimination by chimpanzees (Pan troglodytes): Evidence for approximate numerical representations. Animal Cognition, 11(1), 43-57.
Uller, C., Jaeger, R., Guidry, G., & Martin, C. (2003). Salamanders (Plethodon cinereus) go for more: Rudiments of number in an amphibian. Animal Cognition, 3(2), 105-112.
vanMarle, K., Aw, J., McCrink, K., & Santos, L. R. (2006). How capuchin monkeys (Cebus apella) quantify objects and substances. Journal of Comparative Psychology, 120(4), 416-426.
Vargha-Khadem, F., Gadian, D., Copp, A., & Mishkin, M. (2005). FOXP2 and the neuroanatomy of speech and language. Nature Reviews Neuroscience, 6(2), 131-138.
Vogel, G. (2003). Orangutans, like chimps, heed the cultural call of the collective. Science, 299(5603), 27.
Von Frisch, K. (1967). The Dance Language and Orientation of Bees. Cambridge, MA: Harvard University Press.
Wang, X., Grus, W. E., & Zhang, J. (2006). Gene losses during human origins. PLoS Biology, 4(3), 366-377.
Ward, C. & Smuts, B. B. (2007). Quantity-based judgments in the domestic dog (Canis lupus familiaris). Animal Cognition, 10(1), 71-80.
Wasserman, E. A., Kiedinger, R. E., & Bhatt, R. S. (1988). Conceptual behavior in pigeons: Categories, subcategories, and pseudocategories. Journal of Experimental Psychology: Animal Behavior Processes, 14(3), 235-246.
Weir, A. A. S., Chappell, J., & Kacelnik, A. (2002). Shaping of hooks in New Caledonian crows. Science, 297(5583), 981.
White, D. J., Gros-Louis, J., King, A. P., Papakhian, M. A., & West, M. J. (2007). Constructing culture in cowbirds (Molothrus ater). Journal of Comparative Psychology, 121(2), 113-122.
Wilkinson, A., Chan, H., & Hall, G. (2007). Spatial learning and memory in the tortoise (Geochelone carbonaria). Journal of Comparative Psychology, 121(4), 412-418.
Wilson, E. O. (1975). Sociobiology: The New Synthesis. Cambridge: Harvard University Press.
Wimmer, H., & Perner, J. (1983). Beliefs about beliefs: Representation and constraining function of wrong beliefs in young children’s understanding of deception. Cognition, 13(1), 103-128.
Winter, H., Langbein, L, Krawczak, M., Cooper, D., Jave-Suarez, L. F., Rogers, M. A., et al. (2001). Human type I hair keratin pseudogene hHaA has functional orthologs in the chimpanzee and gorilla: Evidence for recent inactivation of the human gene after the Pan-Homo divergence. Human Genetics, 108(1), 37-42.
Wright, A. A. & Katz, J. (2007). Generalization hypothesis of abstract-concept learning: Learning strategies and related issues in Macaca mulatta, Cebus apella, and Columba livia. Journal of Comparative Psychology, 121(4), 387-397.
Wynn, K. (1992). Addition and subtraction by human infants. Nature, 358, 749-759.
Zentall, T. R. (2002). A cognitive behaviorist approach to the study of animal behavior. The Journal of General Psychology, 129(4), 328-363.
Zucca, P., Milos, N., & Vallortigara, G. (2007). Piagetian object permanence and its development in Eurasian jays (Garrulus glandarius). Animal Cognition, 10(2), 243-258.
Figure Captions
Figure 10-1. Giraffes, which originally had short necks, needed to stretch to reach higher branches, especially in times of food shortage. This stretching would affect the bones and muscles of the neck. According to Lamarck, these acquired characteristics could be passed along to offspring and it was this mechanism that accounted for evolutionary change.
Figure 10-2. A demonstration of natural selection as the mechanism for evolutionary change. According to Darwin, there is variability among animals and the environment determines which of the existing varieties will pass on to the next generation. Over the course of many generations, those varieties conducive to survival are passed along and the length of the giraffe’s neck slowly increases.
Figure 10-3. Gould’s theory of punctuated equilibrium. According to the theory, the pattern of evolutionary change involves occasional spurts of change followed by long periods of relative stability. These spurts of change are in response to drastic environmental changes creating new selective pressures.
Figure 10-4. A simple demonstration of speciation in which groups of fruit flies are separated and an artificial selection procedure is applied based on dietary preferences. After as few as eight generations, the resulting flies will not mate with each other.
Figure 10-5. Animals share many brain structures, however, more phylogenetically advanced species tend to have an increasing amount of gray matter, areas of the cortex that do not have a defined function and are therefore available for higher cognitive processing.
Figure 10-6. The Hebb-Williams test of intelligence for animals.
Figure 10-7. A demonstration of the matching to sample and delayed matching to sample procedures used to test animal memory.
Figure 10-8. Basic procedure used in enrichment studies. Animals are divided into an enriched group which gets social housing and objects to play with and an impoverished group that gets standard laboratory housing. Following a period of exposure to these different environments, various measures are taken to reveal the effects of nurture on “intelligence” including an increased amount of dendritic branching in the enriched group.
Figure 10-9. An open field used in Brown’s (1995) systematic study of the conditions present in a standard enrichment task. Viewed from above and surrounded by plexiglass walls, the open field contained novel objects in the center of each square. Rats were placed in the center square and the number of contacts with novel objects was recorded.
Figure 10-10. Apparatus used by Gagnon and Dore (1994) to test object permanence in dogs. A squeeze toy was initially shown to the dog, then hidden behind one of the boxes. Dogs displayed object permanence when they approached the correct box.
Figure 10-11. The world contains a nearly infinite amount of information that has the potential to enter the memory system, but only those things we attend to will move from the sensory register to the short-term memory. Of the items in short-term memory, only some will be encoded into long-term memory. Once memories are in long term storage, we will eventually need to retrieve them into our working memory. Forgetting can occur at any point in this system.
Figure 10-12. Rats were placed on an FI 20 s schedule (solid dots) or an FI 40 s schedule (open dots). On occasional trials no reinforcement was given at the usual time and the rats’ response rates were recorded. As can be seen, the rats’ peak response rate was at the time they normally were supposed to be receiving food, thus suggesting an ability to measure time intervals.
Figure 10-1
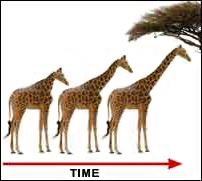
Figure 10-2
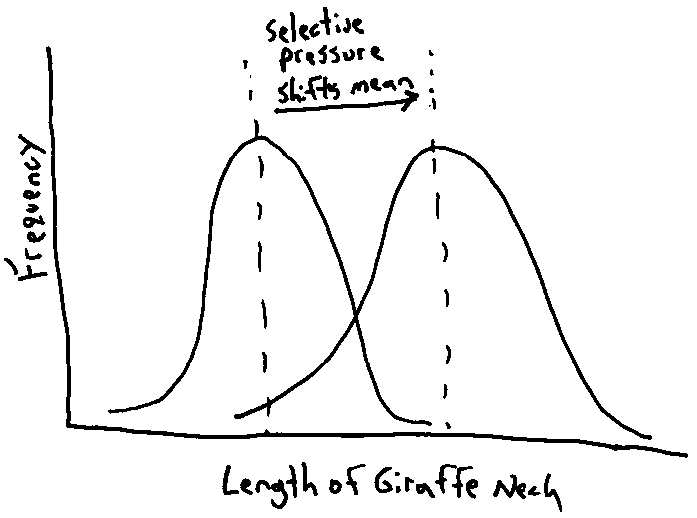
Figure 10-3
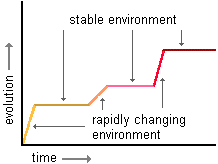
Figure 10-4
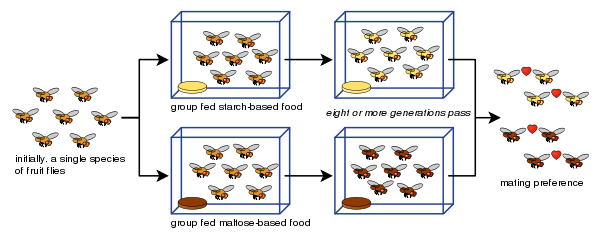
Figure 10-5
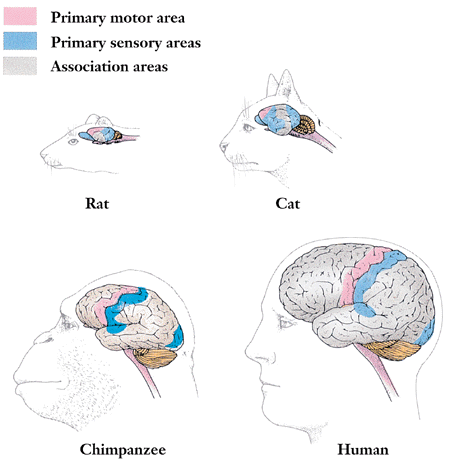
Figure 10-6

Figure 10-7
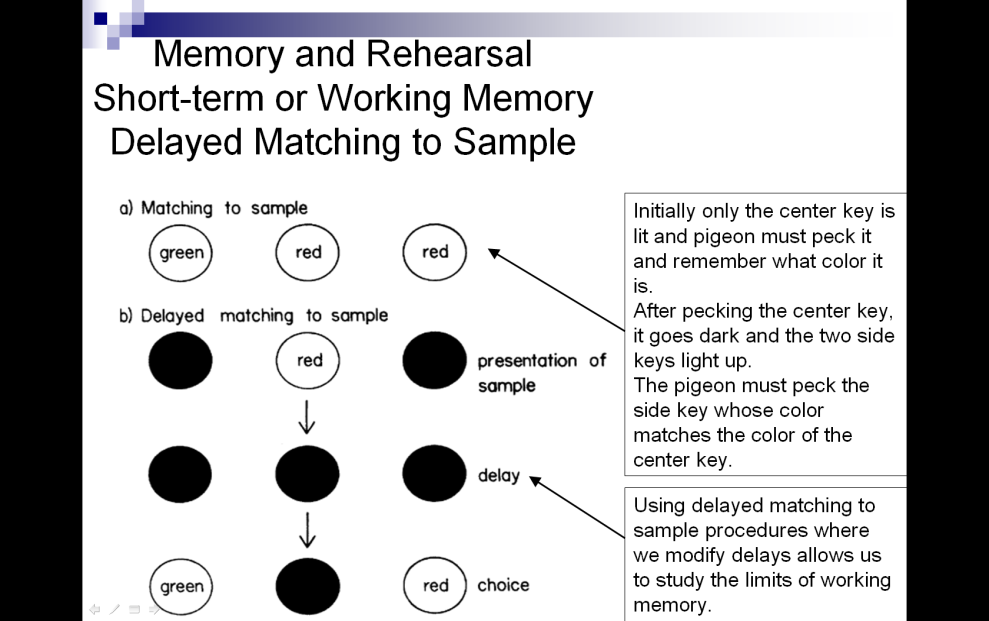
Figure 10-8

Figure 10-9
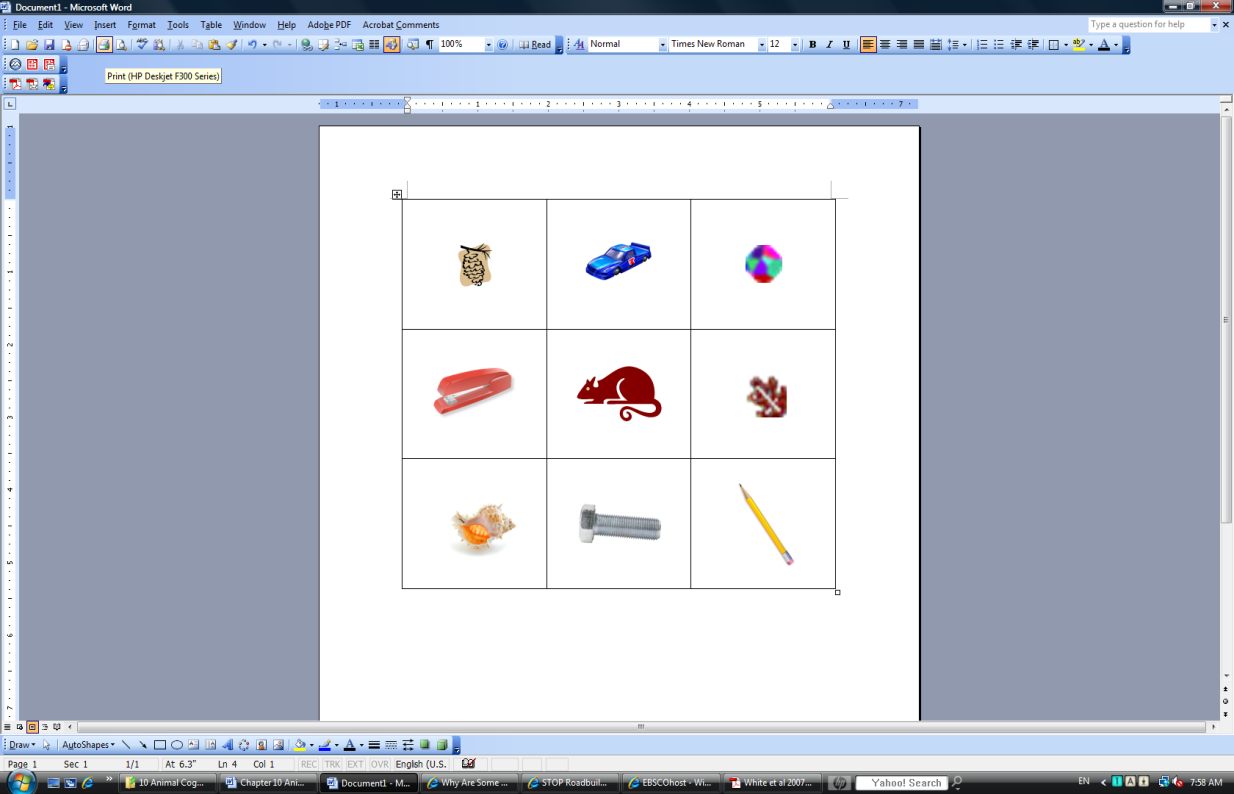
Figure 10-10
Figure 10-11
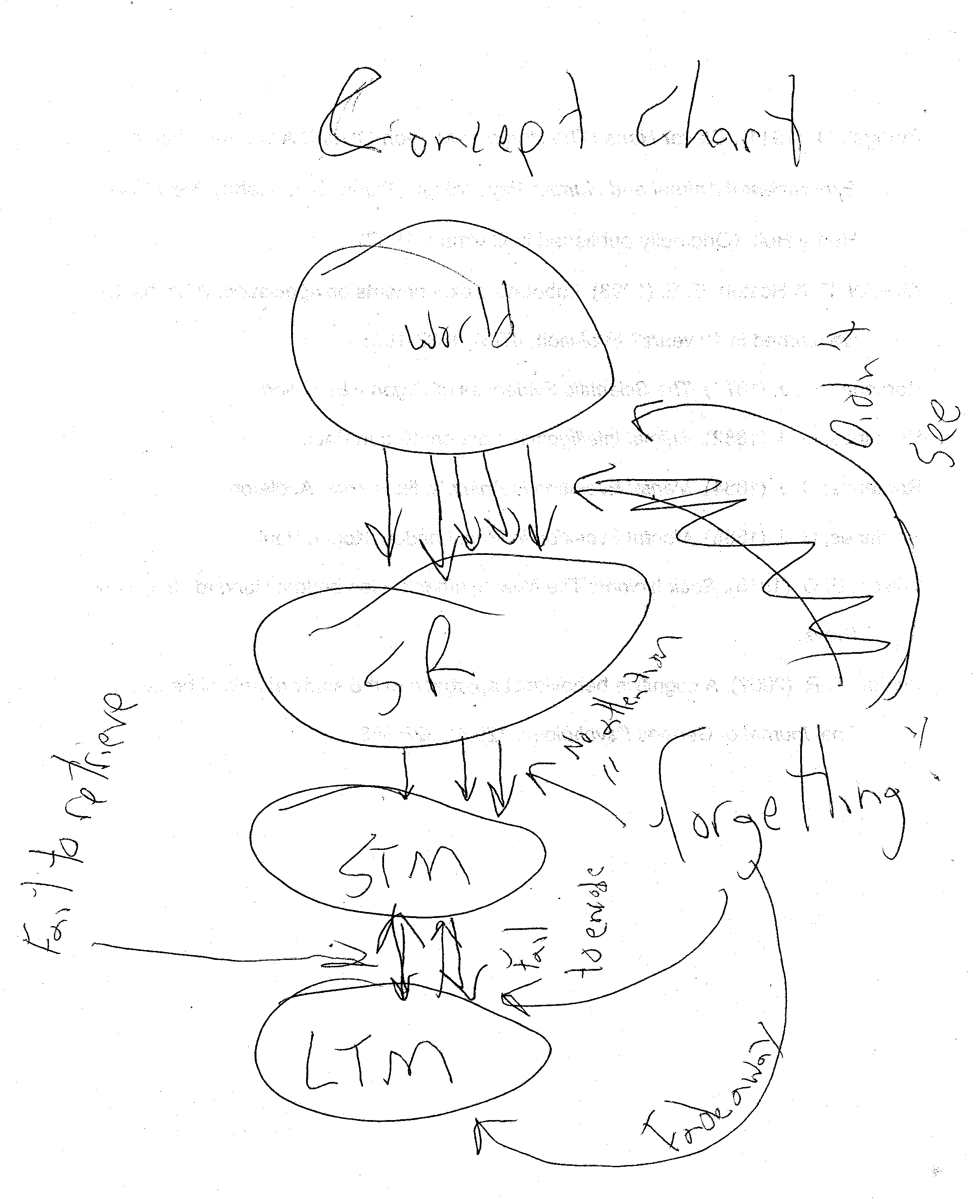
Figure 10-12
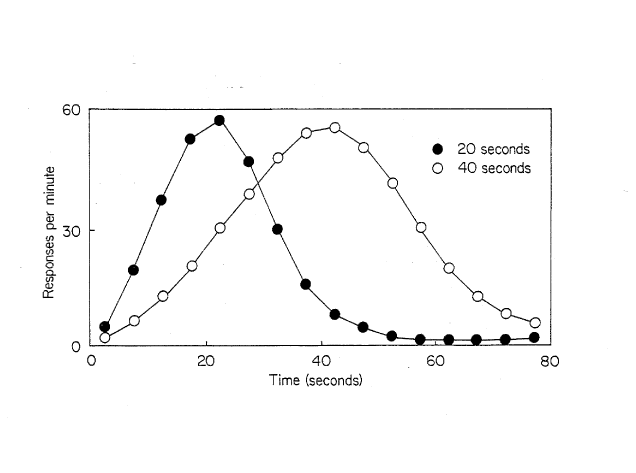
Picture 1

Picture 2

Picture 3

Picture 4

Picture 5

Picture 6

Picture 7
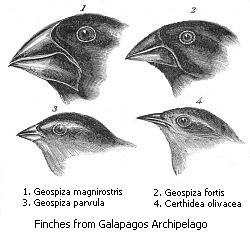
Picture 8

Picture 9
